

 DownLoad:
DownLoad:



 百度学术
百度学术

-
Grinnell[1]最早提出生态位(Niche)概念,用来表示对栖息地再划分的空间单位.自从生态位理论提出后,国外学者首先对其进行了广泛的研究.国内对生态位理论的研究始于20世纪80年代,90年代以后生态位的研究得到广泛关注,成为当前生态学的研究热点之一.生态位宽度和生态位重叠是描述一个物种的生态位及物种间关系的重要数量指标,生态位理论在种间关系、群落结构、物种多样性维持及种群进化的研究中已被广泛应用[2-6],目前的研究主要集中在生态位宽度和生态位重置指标的估算和分析上[7-9].贵州茂兰国家级自然保护区保存着地球同纬度地区原生性较强、分布面积最广的喀斯特森林,是一类特殊的森林生态系统[10],有着丰富的生物多样性,是研究喀斯特森林生态系统物种多样性维持机制的理想场所.前人[11-12]对喀斯特森林生态系统的研究主要集中在植物群落的物种组成、多样性特征和功能群研究等方面,对群落优势种群的生态位特征研究较少[13].本研究对茂兰喀斯特森林不同演替阶段乔木层优势种群生态位宽度和生态位重叠进行分析,揭示茂兰喀斯特森林优势乔木种群的种间关系和更替规律,为退化喀斯特森林生态系统的恢复和重建提供理论参考.
全文HTML
-
研究区位于贵州省南部贵州、广西交界处的茂兰国家级自然保护区(25°09′-25°20′N,107°52′-108°05′E),总面积约20 000 hm2,东西宽22.8 km,南北长21.8 km.研究区为典型的喀斯特峰丛地貌,最高海拔1 078.6 m,最低430 m,平均在800 m以上.气象站海拔806 m,当地年均温15.3 ℃,7月均温26.4 ℃,1月均温8.3 ℃,≥10 ℃积温5 727.9 ℃,年均降雨量1 320.5 mm.全年平均相对湿度83%.全区除局部地点覆盖有少量砂页岩外,其余区域主要为由纯质石灰岩及白云岩构成的典型喀斯特峰丛地貌,属于裸露型喀斯特,被称作“典型的喀斯特生境”.与生长在这种生境基质上的森林融合在一起,构成了中亚热带地区独特的喀斯特森林自然综合体[10].
-
本研究采用以空间代替时间的研究方法,分别在3个不同演替阶段(演替10年为早期、演替20年为中期、演替40年为晚期)的典型地段设置面积10 m×10 m的样地10个,共设置30个样地.沿每个样地的对角线设置5 m×5 m大小的基本样方,记载每个样方内乔木的种类、个体数、胸径、高度、冠幅等内容.
-
1) 生态位宽度
生态位宽度(B)是反映物种对资源利用程度的参数.采用Levins提出的公式[9]:
式中:B为某个种生态位宽度;S为资源单位数;Pij是种i对j资源利用率或它在该资源状态上的分布比例量.
2) 生态位重叠
生态位重叠是指一定资源序列上,2个物种利用同等级资源而相互重叠的情况,采用相似性比例或比例重叠公式:
式中:Cih为物种i与物种h生态位重叠;S为资源单位数;Pij与Phj分别为物种i与物种h在第j个资源单位中所占的比例.
3) 依据公式IV=(相对密度+相对频度+相对显著度)/3计算物种重要值
1.1. 研究区概况
1.2. 研究方法
1.2.1. 野外调查
1.2.2. 数据处理
-
在所调查的30个样地中,乔木层共记录植物64种,分属36科54属.为更好地探讨物种在群落中的地位和作用,本研究计算了各演替阶段乔木层物种重要值和变异系数.由表 1可知,演替早期重要值最大的是轮叶木姜子(76.77),其次是巴东荚蒾(42.08),而变异系数最大的是黄梨木(158.57%),其次是香叶树(152.73),两者的重要值均较小;演替中期重要值最大的是轮叶木姜子,其值较演替早期有所降低(32.52),其次是四照花(30.66),在该演替阶段变异系数最大的是轮叶木姜子和云贵鹅耳枥,其值分别为184.86%和91.10%;演替后期重要值最大的仍然是轮叶木姜子(48.00),可见其在整个演替序列中是优势种以及群落的建群种,其次是青冈栎(29.91),该演替阶段变异系数最大的几个种群变异差异较小,大致分布在88%~89%内.
-
从表 2可知,在不同演替阶段主要物种的生态位总宽度排序从大到小依次为樟叶槭、革叶铁榄、翅荚香槐、石岩枫、齿叶黄皮、皱叶海桐、狭叶润楠,且其生态位宽度值大于2.演替早期,樟叶槭的生态位宽度最大,为1.02,其他生态位较宽的物种还有皱叶海桐、翅荚香槐、革叶铁榄、柿树、巴东荚蒾、齿叶黄皮、轮叶木姜子,分别为0.99,0.96,0.94,0.90,0.89,0.89,0.87.表明樟叶槭等8种树种在演替早期分布范围较广,对环境资源综合利用能力强,具有较强的适应能力;演替中期,樟叶槭、翅荚香槐等物种的生态位宽度有所下降,石岩枫的生态位宽度提高到该演替阶段的最大值,为0.91,灯台树和大叶冬青的生态位宽度有较大提升,分别从0.20和0.42提高到了0.46和0.53,该阶段青桐开始出现,但生态位宽度值较小,仅为0.30;演替晚期,齿叶黄皮的生态位宽度最大,为0.76,巴东荚蒾、樟叶槭、石岩枫和革叶铁榄的生态位宽度略有下降,此阶段出现的新物种中生态位宽度最大的是鹅耳枥,其值为0.56,其余新物种生态位宽度值大约在0.09~0.45之间.可见,在群落演替的过程中,随着生境条件的改变,一些物种的生态位宽度值也随之发生改变,从而导致了它们在整个演替过程中的动态变化.
-
由表 3可知,演替早期阶段,生态位重叠值较大的几个种对分别是青冈栎-黄梨木(0.97)、润楠-鹅耳枥(0.92)、四照花-香叶树(0.91)、皱叶海桐-翅荚香槐(0.88)、润楠-翅荚香槐(0.88)、香港四照花-椤木石楠(0.88),表明它们在同一资源位上出现的频率较多,生态相似性较大.总体来看,青冈栎、皱叶海桐、翅荚香槐等树种与其他树种的生态位重叠程度较高.在此阶段,重叠值大于0.70的种对有55对,占总对数的16%,其他的树种中有很多物种种对重叠值为0或很小,表明演替早期阶段森林中因利用相同的资源位而产生的种间竞争不激烈.
-
由表 4可知,演替中期阶段,生态位重叠值较大的几个种对分别是革叶铁榄-石岩枫(0.98)、狭叶润楠-樟叶槭(0.98)、狭叶润楠-石岩枫(0.97)、香港四照花-皱叶海桐(0.97)、樟叶槭-皱叶海桐(0.95).石岩枫、樟叶槭、皱叶海桐等种群间生态位重叠较高,这是因为优势种石岩枫与这些种群均具有较宽的生态位,在群落的分布上较多重叠与混生,表明种群间的生态位重叠分离明显.而另有某些种群如润楠,本身生态位很小,与其它多数种群间的生态位重叠值非常低,有的甚至为0,表明这些物种在这个阶段的生境里生态幅窄小,对资源利用能力较差,仅为群落的偶见种.
-
演替晚期阶段,作为3个演替阶段中生态位重叠均值最高的一个阶段,其资源利用性竞争较前2个阶段激烈.由表 5可知,生态位重叠值高达0.99的就有4个种对,其中生态位重叠值大于0.70的种对共有98对,占整个演替阶段乔木种群的33%,较演替早期提升了大约一倍.优势种齿叶黄皮与石岩枫、皱叶海桐的生态位重叠值达0.91和0.90,在演替晚期阶段优势种群与其它种群间生态位重叠均较高,表明群落中的大多数种群在资源利用上具有极大的相似性.
-
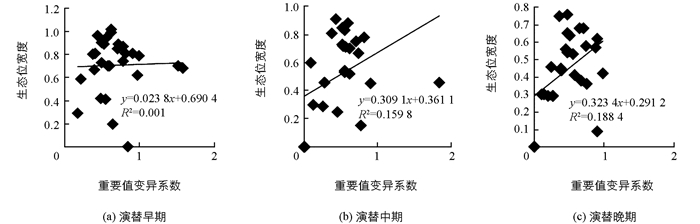
分别将各演替阶段乔木层优势种生态位宽度与重要值和重要值变异系数进行线性回归.由图 1可知,3个演替阶段种群的生态位宽度与重要值呈正相关关系,但相关性极不显著,其相关系数分别为0.355 6,0.462 1,0.177 8.由图 2可知,各演替阶段种群生态位宽度与重要值变异系数也呈正相关关系,其相关系数分别为0.001,0.159 8,0.188 4,说明各演替阶段种群生态位宽度随重要值和重要值变异系数的增大而增大.
2.1. 不同演替阶段物种重要值
2.2. 生态位宽度
2.3. 生态位重叠
2.3.1. 演替早期
2.3.2. 演替中期
2.3.3. 演替晚期
2.4. 生态位宽度与重要值及重要值变异系数的关系
-
生态位分化是竞争物种共存的基础,是物种进化的动力,也是种群结构变化和种群演替的主要原因之一[2].在茂兰喀斯特森林中,樟叶槭、石岩枫、齿叶黄皮、革叶铁榄、狭叶润楠、柿树、轮叶木姜子等种群,数量多,分布广,在群落内部环境中起重要作用,它们生态幅较大,竞争能力较强,处于优势种地位,所以它们的生态位宽度也较大.相反,川桂、山矾、润楠等种群分布较为集中或数量较少,利用环境资源的能力较弱,在群落中属于伴生种,对该区生态环境的适应能力次于优势种.不同物种生态位的差异,说明它们的资源利用能力不同,在不同资源空间中的生态适应性也不同.一般而言,生态位宽度大的种群由于利用资源能力较强、分布较广,生态位重叠较大.生态位宽度小的种群由于对资源的利用能力较弱、分布范围小而与其它种群间的生态位重叠较小.而本研究发现,在乔木种群的整个演替过程中,生态位重叠均值是呈上升趋势,演替早期最低,演替晚期最高.随着演替的进展,各物种出现了一定程度的生态位分化,生态位宽度呈下降趋势,而生态位重叠均值却呈上升趋势,说明生态位宽度与生态位重叠没有相关性,生态位宽度大或者小的种群,它所构成的种对间生态位重叠既有大的也有小的.如演替晚期的优势种齿叶黄皮尽管生态位较宽,但与大多数种群间生态位重叠并不高,这可能是齿叶黄皮竞争力较强排斥其它物种与其共生和分享资源的缘故,而苦木和灯台树的生态位宽度很小,但其生态位重叠值却不低.可见在群落演变中,植物对环境资源存在着较为激烈的竞争和较高的空间异质性,这与刘巍等[14]对长白山冷杉群落主要种群生态位特征的研究结果一致,其次,还与种群的生物学特性的差异密切相关.演替早期阶段,最大重叠值的种对青冈栎-黄梨木其生态位宽度值不大,这是由于此阶段这些乔木种群还处于幼苗阶段,盖度低,竞争小,群落组成结构还不够稳定.随着演替的进展,中期阶段群落环境趋于稳定,大量的树种开始出现种间竞争,如狭叶润楠-樟叶槭的生态位宽度较小,而生态位重叠值很大,这是由于狭叶润楠是耐荫树种,生于湿润阴坡山谷,而樟叶槭是喜阳树种,耐干燥瘠薄,它们之间由于对资源利用方式的差异以及对限制因子的耐受程度的不同致使生态位发生分异.演替晚期阶段,群落已趋于稳定,在此阶段中,多脉榆、翅荚香槐、圆叶乌桕等乔木都为阳性树种,且分布在同一林层,对光照、水分、土壤养分等环境资源的需求相似,故它们之间的一些种对重叠值都很大.这种现象出现在演替的各个阶段中,充分说明生态位宽度与生态位重叠之间的关系复杂,除与树种的生物学特性有关外,还与喀斯特森林生境的不均匀性和异质性有关.对茂兰喀斯特森林不同演替阶段优势乔木种群的生态位宽度和生态位重叠的研究,能较好地解释群落演替过程中的种间关系及物种对环境的响应等生态过程的演变规律,提高对植物在群落中的地位和作用的认识,加强植被的保护、修复和重建.