


 下载:
下载:
-
环境变化对植物的生长和生态系统结构与功能产生重要影响,而植物也通过自我调节响应环境,从而影响生态系统[1].其中,功能性状与资源获取、利用和保存能力有关,从而对植物的定居、生存和适应有着潜在重要影响,其属性能够单独或联合指示植物对环境变化的响应,对研究植物对环境变化的响应及生态系统的变化具有重要的作用[2-3].
功能性状受多种环境因子的影响.例如,叶片大小和比叶面积随着光照的增强而明显减小[4].叶片的氮、磷含量随年均温和降水量的降低有上升趋势,而叶片大小及氮磷比降低[5].此外,随着海拔的升高,叶干质量和叶面积降低,从而获取较高的抗冻性[6].也就是说,物种通过性状的可塑性及遗传多样性来适应不同的环境,这对于研究植物对环境变化的响应机制有重要作用[7].
另一方面,不同功能群植物往往具有不同的功能性状.落叶植物具有较高的比叶面积、光照利用效率、较短的叶寿命和较大的叶片,从而在短时间内积累足够的光合作用产物[8];相反,常绿植物比叶面积、光照利用效率较低,同时具有较小的叶片和较长的叶寿命[9].植株高度同样影响功能性状,灌木具有较厚的叶片,高的木材密度、导管密度和细的导管直径,同时生长速率较慢而存活率较高[10-11].
目前,许多研究围绕植物功能性状对海拔变化的响应进行了研究[12-13],但在乡—郊—城梯度上植物性状的可塑性研究较少,尤其是不同发育阶段功能性状的差异,这对于进化背景和资源分布异质的环境下植物可塑性的研究意义重大[14].因此,本研究以重庆市14种木本植物为研究对象,测定其枝干、叶片、花和果实性状,研究不同功能群、生境类型和发育阶段下木本植物营养器官如茎和叶和繁殖器官包括花、果实及种子的差异性,试图探明功能群、生境变化和发育阶段对功能性状差异的影响及功能性状对不同生境的适应机制,同时探讨哪些环境因子对功能性状差异产生了影响,为植物进化及城市园林植物配置提供依据.
全文HTML
-
研究区域为重庆市主城区(28°27′-30°26′ N、105°17′-107°27′ E),平均海拔400 m,地形地貌以丘陵山地为主.气候类型为中亚热带湿润季风型气候,年均降水量在1 000~1 200 mm之间,空气湿度大.采样区域分布在渝中区、江北区、南岸区、九龙坡区、沙坪坝区、北碚区和渝北区,依据绿地覆盖率、热岛强度及人为干扰强度将采样地点划分为森林公园、郊区公园和城市公园3种生境.森林公园包括南山植物园、歌乐山森林公园、中央公园、园博园和缙云山;郊区公园包括西南大学、重庆大学、照母山森林公园和鸿恩寺森林公园;城市公园包括沙坪坝公园、动物园、嘉陵公园、百林公园、歌乐山文化广场和鹅岭公园.其中一些校园、广场等绿地覆盖率相对较高,也将其作为公园类型.
-
本研究采集的枝条为:成熟、健康且末端不分枝的当年生枝条、着生完全开放花朵和花蕾的当年生枝条及着生成熟果实的当年生枝条,包括其上的附属物(叶、茎、花和果实).实际采样中,部分植物的花朵和果实着生在二年生枝条上,也将其包含在本次研究中.实验于2017年9月-2018年9月进行,根据植物物候的差异,分别在植物完成展叶及枝条生长时、花期和果熟期,于远离样地边缘处,每个物种随机选择3~6株成熟个体,在每株植物外冠层东西南北4个方向随机采集4个没有明显叶、花和果实损伤的枝条,采集后立即放入自封袋,带回实验室测量相关指标.
-
首先测量带回的实验材料的小枝茎长、茎直径,并记录叶片数量,之后放在恒温鼓风干燥箱中,在70 ℃下条件下烘干48 h后至恒质量,用FA1004万分之一电子天平测量叶、茎、花、果实等干质量指标.对于性状原始参数测量方法如下:小枝茎长、茎直径利用直尺或游标卡尺测得;茎干质量(SM)、叶干质量(LM)、花干质量(FLM)、花被干质量(PM)、雄蕊与雌蕊群干质量(PIM)、果实干质量(FM)、种子干质量(SEM)利用电子天平测量烘干后质量;叶面积利用透明玻璃盖板压平叶片并采集电子照片,利用Adobe Photoshop CS5和AutoCAD 2010软件计算.计算参数包括:茎截面积(TCSA,茎直径×茎直径×π/4)、茎密度(TD,茎干质量/(小枝茎长×茎截面积))、单叶面积(ILA,叶面积/叶片数量)、单叶干质量(ILM,叶干质量/叶片数量)、比叶面积(SLA,叶面积/叶干质量).
-
采用Kestrel 4000手持式风速仪测定风速(Wis)、温度(Tem)、湿度(Hum),GPS手持仪测定海拔(Ele),便携式氮氧化物含量检测仪测定氮氧化物(NOx),手持DUSTMATE粉尘检测仪测定TSP和PM 2.5.测定于2018年3月10日、6月10日、9月10日的10:00-15:00进行,对样地内18~25个样点进行观测,每个样点选择5个观测点,观测点距地面约1.5 m,于第2天进行重复实验.
下垫面性质测定方法:利用Google Earth v7.3.0截取采样地点的卫星图片,导入AutoCAD 2010软件勾勒出采样地点的建筑、硬质铺装、植被覆盖及整个区域,并将其生成面域,利用软件本身矢量数据计算功能得到相应的面积,最后利用公式(相应指标面积/采样地点面积×100)计算出建筑面积占比(Bar)、硬质铺装占比(Hpr)及植被覆盖度(Vec)(表 2).
-
在进行数据分析之前,为保证数据的正态分布性,对各性状参数进行对数处理.利用IBM SPSS Statistics 20.0软件对性状参数进行方差分析和主成分分析,用新复极差法进行多重比较;利用CANOCO 4.5软件绘制物种在性状上的分布的PCA排序图,对环境因子和性状进行冗余分析并绘制RDA排序图;利用主成分分析中的载荷值、特征值及标准化的性状参数建立主成分方程,实现对研究对象的综合评价;利用一般线性模型(GLM)剖析生境类型和功能群对性状的相对影响.除排序图外均采用Origin 9.1软件作图.
1.1. 研究区域
1.2. 样品采集
1.3. 指标测定
1.3.1. 功能性状指标测定
1.3.2. 环境因子指标测定
1.4. 数据分析
-
除TD和SLA外,SM,TCSA,ILA及ILM在不同物种间差异极具有统计学意义(p<0.01);不同发育阶段物种间茎性状差异具有统计学意义(p<0.05),叶性状中除SLA外,ILA及ILM差异极具有统计学意义(p<0.01).另外,2种因素的交互作用对于SM,ILA,ILM和SLA的影响达到极具有统计学意义水平(p<0.01),对TCSA和TD影响不具有统计学意义(表 3).
-
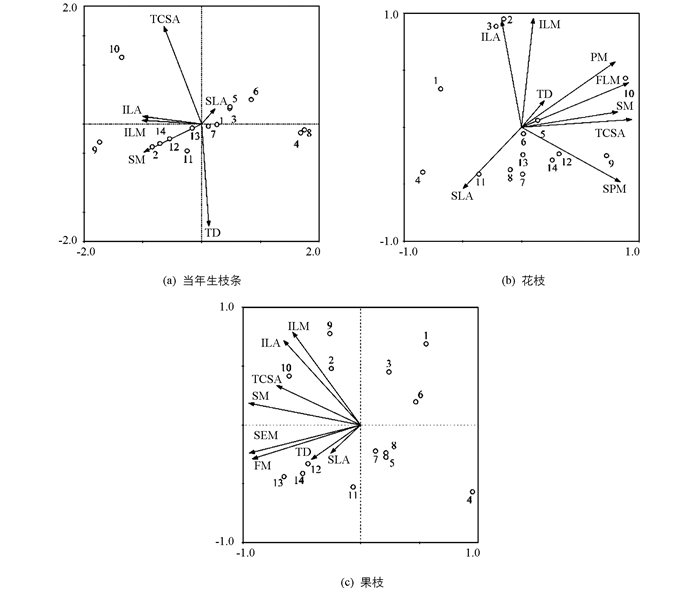
在营养器官中,紫荆、玉兰的SM和TCSA最大,红花檵木最小;桃的TD最大,樟最小;玉兰、天竺桂的ILA和ILM最大,红花檵木最小;紫叶李的SLA最大,玉兰最小.在繁殖器官中,紫荆、玉兰的FLM、PM及SPM最大,红花檵木最小;玉兰的FM和SEM最大,樟最小.此外,主成分分析结果表明功能性状间关系十分密切.茎性状中SM与TCSA、TD在花枝和果枝中相关具有统计学意义;叶性状中ILA和ILM正相关,而与SLA负相关;花性状中PM与FLM正相关具有统计学意义,与SPM相关性减弱;果性状中,FM与SEM正相关具有统计学意义(图 1).
-
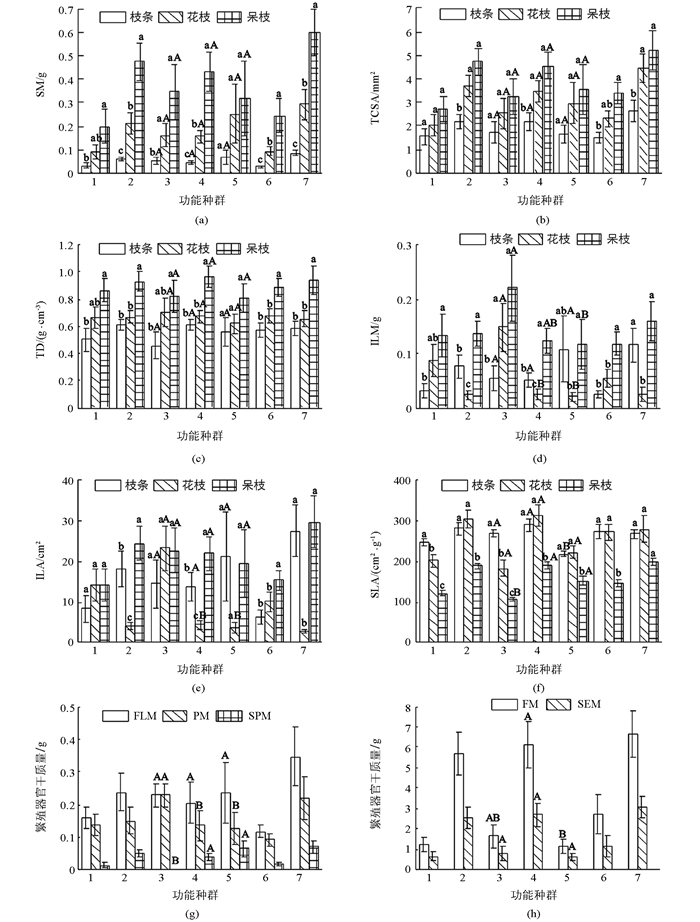
在生活型上,功能性状在当年生枝条上差异不具有统计学意义,而花枝的SM,TCSA,ILM,ILA和SPM及果枝的SM,TCSA,ILA,SLA,FM和SM差异具有统计学意义(p<0.05),且落叶大于常绿植物(花枝的ILA和ILM除外);在生长型上,仅有当年生枝条的SLA、花枝的ILA,ILM和SPM及果枝的SLA,FM差异具有统计学意义(p<0.05),且乔木大于灌木;在开花习性上,当年生枝条的TD和SLA、花枝的ILM和SLA及果枝的ILM差异不具有统计学意义,其余性状差异均具有统计学意义(p<0.05).花枝的ILM,ILA和SLA表现为先叶后花大于先花后叶植物,其余枝条和性状相反(图 2).
-
在生活型上,除常绿植物的TCSA和ILA外,其余性状差异具有统计学意义(p<0.05).在生长型上,乔木的SM,ILM和SLA、小乔木的SM,TCSA,TD,ILA,ILM和SLA及灌木的SLA差异具有统计学意义(p<0.05).在开花习性上,除先花后叶植物的SLA外,其余性状均差异具有统计学意义(p<0.05).具体表现为不同功能群植物的SM,TCSA,TD,ILA和ILM逐渐增大,SLA相反(图 2).
-
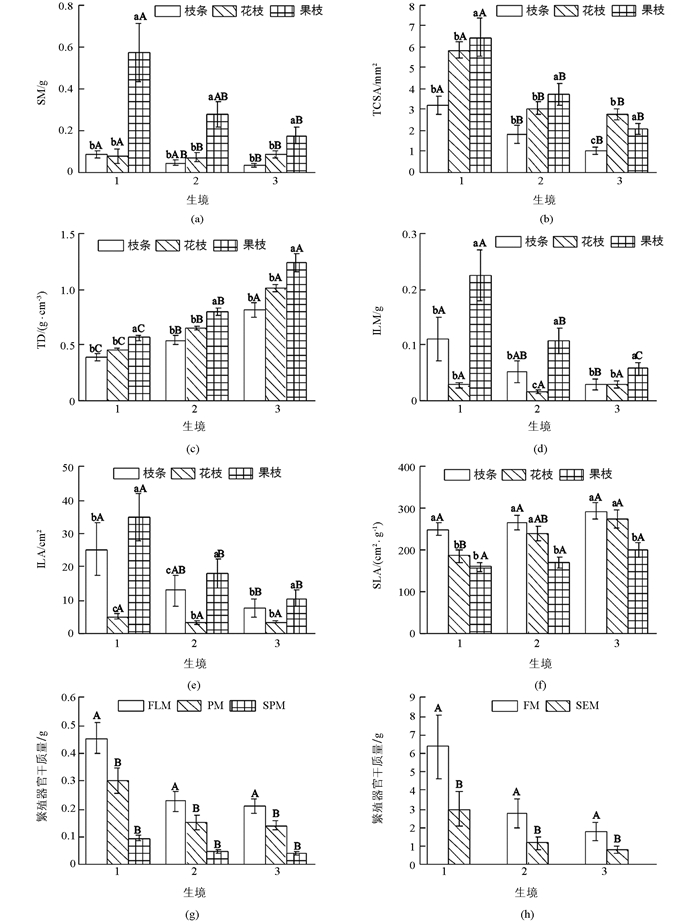
除SLA、花枝的ILA和ILM外,其余功能性状在生境下差异具有统计学意义(p<0.05),表现为茎性状中TD随乡—郊—城梯度变化逐渐增大,而SM和TCSA则相反(图 3(a)-图 3(c));叶性状中ILA和ILM逐渐下降(花枝为先下降后升高),而SLA有增大的趋势(图 3(d)-图 3(f));繁殖器官中FLM,PM,SPM,SEM和FM均表现为下降的趋势(图 3(g)-图 3(h)).
-
不同发育阶段物种的功能性状差异具有统计学意义(p<0.05).除SLA外,其余性状参数在果枝中均最高.其中,SM,TCSA和TD随着新—花—果梯度变化表现为升高的趋势,而ILA和ILM在花枝中最小,SLA随梯度变化逐渐降低(图 3).
-
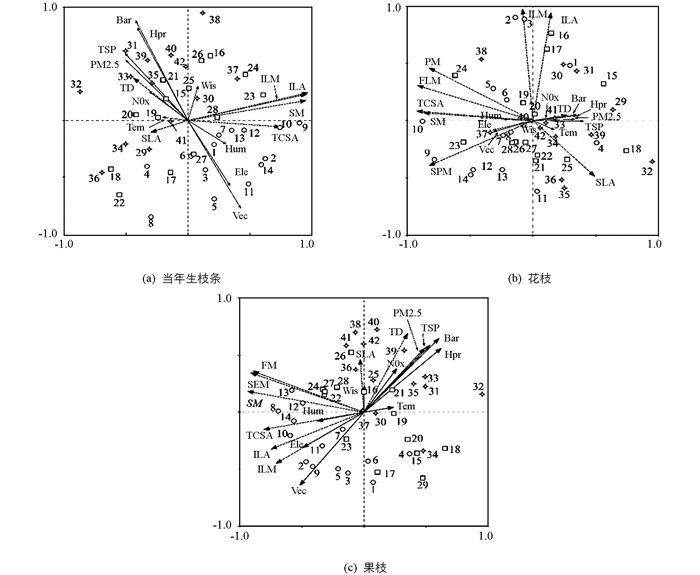
对环境因子和功能性状进行冗余分析,结果表明,Tem,NOx,TSP,PM2.5,Bar和Hpr正相关具有统计学意义,与Hum,Ele,Vec负相关具有统计学意义,其中Hum,Ele,Vec与性状参数TD和SLA负相关,与其他性状正相关.在与植物分布的关系中,城市环境温度和污染较高,以至于在城市中生长的植物功能性状受到戕害,而森林环境与植物生长的自然生境最为接近,植物具有最大的SM,ILA,ILM,FLM及FM(图 4).
-
结果表明,功能性状受到功能群、环境和发育阶段的共同影响.利用广义线性模型进一步定量分析各影响因素的对功能性状的影响,结果表明,环境因子对当年生枝条和果枝性状变异解释的比例最高,分别为9.9%~52.9%和48.7%~149.4%,除SLA外对其他性状变异的影响均具有统计学意义;生长型是花枝性状变异的主要来源,解释的比例为1.0%~50.0%;在不同发育阶段中,发育阶段和环境对于性状变异的影响均具有统计学意义,是变异的主要来源,解释的比例分别为23.4%~58.8%和5.4%~62.7%(表 4).
2.1. 物种功能性状总体差异特征
2.1.1. 不同物种功能性状的差异
2.1.2. 物种在功能性状上的分布
2.2. 不同功能群间功能性状的差异
2.2.1. 功能群在同一发育阶段功能性状的差异
2.2.2. 功能群在不同发育阶段功能性状的差异
2.3. 不同生境下功能性状的差异
2.3.1. 不同生境下同一发育阶段功能性状的差异
2.3.2. 不同生境下不同发育阶段功能性状的差异
2.3.3. 环境因子对功能性状的影响及综合评价
2.4. 功能群和生境类型对功能性状的综合影响
-
比叶面积和叶干物质含量在一定程度上反映了植物对资源获取和利用的能力[15-16].常绿植物的比叶面积较低,叶片内物质主要用于构建保卫组织或增加叶肉细胞密度,叶片面积较小,叶寿命较长;落叶植物单位质量的叶片面积较大,叶氮含量、光捕获量和光合能力均较高[17].本研究中,落叶植物的比叶面积显著大于常绿植物,表明了常绿植物倾向于延长叶寿命而落叶植物通过提高光合速率以保证积累足够的光合产物;叶干物质含量在花枝中表现为常绿大于落叶植物,可能是由于落叶植物中紫荆、桃属于先花后叶植物,在开花的同时叶片还未长出或正在萌发.此外,落叶植物的茎干质量和茎密度也大于常绿植物,即落叶植物其机械支撑能力较强,对病原体抵抗能力强[18].
植株高度决定了空间光照和资源的分布不均,同时可以反映植物对于环境的不同适应机制[11].乔木在短时间内通过提高光合速率来偿还构成叶片的成本,其比叶面积和叶干物质含量较高[19].本研究中,乔木的叶、茎、花和果实的含量均高于灌木,可能由于生态位差异,乔木拦截光照和光合作用能力较强,使得光合作用同化的产物增多,对于植物各器官的投入也增多;同时枝条除了供给叶片生长需要的水分和养分外,还需提供支撑作用,叶片、花和果实越大,所需要的支撑作用越大,对于小枝的投入也越多[20].
-
茎具有支撑、保存水分、养分及传导树液体的作用,其中,茎木质密度反映了木材内细胞壁积累物质的能力[8];叶片作为植物光合作用的主要器官,由于其属于扁平结构与环境接触的面积大,对于环境变化非常敏感[21].高温、干旱使得植物的叶寿命和比叶面积降低,以降低植物蒸腾作用丧失的水分[8].本研究中,城市公园中茎干质量、茎截面积和叶干物质含量显著低于郊区公园和森林公园,茎密度在3种生境下变化趋势相反,说明了城市环境使得植物的营养器官受到一定戕害,而植物一方面通过降低与外界的接触面积来降低污染对其造成的伤害;另一方面,通过增大茎密度以在更干及营养供应不足的土壤中或更强的物理干扰条件下生长[18].此外,植物为了防止蒸腾作用过于强烈、城市热岛高温损伤,需要有一部分物质构建角质膜等保卫结构,使得叶片光合累积同化产物的能力下降,形成小叶片[22].
花、果实和种子是植物繁衍的重要器官,对基因延续和保护物种意义重大[23].随海拔的升高,花粉数逐渐降低,而花柱、花丝、花大小和种子大小逐渐增大[24-25].本研究中,城市公园中花和果实质量均趋于降低,即植物投入到繁殖器官的能量减少以至于植物的繁殖受到戕害,从而推知城市植物花、果实和种子减小将导致城市植物种子和幼苗竞争力下降从而影响植物的自然更新. RDA排序图很好地表明了功能性状与环境因子之间的关系,温度和污染是主控因子.在污染程度高的区域,植物通过减小花的大小降低对水分、养分等资源的消耗,将多余的营养和能量分配给其他器官或转移到合成花气味或色素的物种中,以增强资源竞争的能力或吸引传粉者[26].此外,城市温度较高,植物生长季延长,植物采用“每天开少量花,延长开花时间”的策略提高繁殖成功率;反之,森林环境中生长季缩短,植物需要增大花、果实和种子构件的光合产物投入,尤其在大种子内部可以储存较多的能量以供植物幼苗生长过程所需的营养物质[27].
-
本研究中,果枝中茎干质量、茎密度、叶干物质含量和叶面积均显著大于花枝和新生枝,而比叶面积则相反,这体现了3种发育阶段的物种本身所具有的差异:新生枝需要更强的光合作用来保持自身的生长,从而需要较高的比叶面积提高光合速率,而果枝叶片内的物质需要有一部分用于构建保卫结构、增加叶肉细胞密度,延长叶寿命,因此比叶面积较低[28].随着发育历程的变化,生活在相似资源和环境中的植物均通过增大叶片和茎的干物质储存来适应环境的改变,即由于环境过滤器作用使得植物功能策略趋同[29].
综上可知,功能群、生境类型和发育阶段对于功能性状差异均有影响.城市环境对植物功能性状总体有显著的负面效应,在各环境因子中,温度、硬质铺装占比及PM 2.5对功能性状影响较为显著,而植物通过不同功能性状的调节,以适应高温、污染及干旱的特殊环境.因此,为缓解城市环境对园林树木生长的不利影响,应参考叶干物质含量及茎密度等指标筛选耐旱、耐热和耐污染的树种.诚然,单独的功能性状仅仅能够揭示出植物对环境梯度响应一个方面,而多个性状的组合及与生物量相互联系可能更好地反映植物对环境梯度或适应对策的全貌.因此,在研究植物与环境关系时,应尽可能地观测多个性状及考虑它们之间的相互关系,从而为进化机制与园林植物应用提供一定的基础.