


 下载:
下载:
-
生物量是植物功能性状之一,反映了生态系统生产力的大小,是生态系统物质循环和能量流动的体现[1],其实质是绿色植物通过同化器官的光合作用在单位面积内积累有机质和能量.如何有效地分配生物量对于物种的生存和繁殖有巨大的作用[2],平衡生长假说[3]认为,植物在响应环境时,通过调节不同器官中的生物量分配比例,以将资源分配到最需要的器官中,从而更有效地获取水分、养分和光照等受限的资源.即当某一构件获得收益时,另一构件就需要付出适当的代价;此外,Mccarthy和Enquist[4]发现资源匮乏时,植物叶生物量占总生物量的3/4,而根生物量和茎生物量各占1/8,即植物生物量分配在各个组织器官之间有某种平衡.确定和解释这些构件的相关性,有助于理解物种对环境的响应、物种共存和生态系统的稳定性.
植物不同构件间的权衡关系为异速生长是普遍承认的规律[5].其中,叶片通过光合作用进行物质生产,茎具有物理支持、保存水分和养分及传导树液体的功能,花、果实和种子对于植物繁衍、基因延续和保护物种起着至关重要的作用. Corner[6]法则认为枝条越粗,其上生长的附属物——叶、花和果也较大,那么可以推测在小枝茎基础上叶、花和果之间也存在正相关的关系.同时,这种相关性受到温度、光照和水分等因素的影响.在低的土壤养分、低降雨量、低温和高的海拔中,在小枝水平上,植物通常分配更多的生物量给茎,并且具有更高的比叶质量[7].张艳茹等[8]研究了低海拔的21种常绿杜鹃,结果显示,总果实干质量和种子千粒干质量随着总叶片干质量及总叶面积的增加而增加,即叶片大小对果实的生物量和种子千粒干质量的影响具有统计学意义.
目前,许多研究围绕生物量在不同生活型及植物各器官之间的相关性进行,但在乡—郊—城梯度上生物量差异及植物各器官之间的相关性研究较少,尤其在不同发育阶段生物量及相关性差异的研究更是鲜见,而这方面的研究对于植物在不同环境下的可塑性研究意义重大.本研究选取重庆主城区常见的14种木本植物,以新生期、花期和果期的当年新生枝条为研究对象,测定枝干、叶片、花和果实性状,研究不同功能群、生境类型和发育阶段的生物量分配及植物各器官之间的差异,试图探明功能群、生境变化和发育阶段对生物量分配及功能性状相关性差异的影响,为植物进化及城市园林植物配置提供依据.
全文HTML
-
研究区域为重庆主城区(28°27′-30°26′ N、105°17′-107°27′ E),平均海拔为400 m,地势高低起伏,地形地貌以丘陵山地为主.气候类型为中亚热带湿润季风型气候,年均降水量在1 000~1 200 mm之间,空气湿度大.其中采样区域分为3种生境类型(表 1).
-
以成熟、健康且末端不分枝的当年新生枝条、着生完全开放花朵的当年新生枝条及着生成熟果实的当年新生枝条为研究对象,包括其上的附属物(叶、茎、花和果实).实际采样中,部分植物的花朵和果实着生在二年生枝条上,也将其包含在本次研究中(表 2).实验于2017年10月-2018年10月进行,根据植物物候的差异,分别在每个物种完成展叶及枝条生长时、花期、果熟期,随机选择3~6株成熟个体,在植物外冠层东西南北4个方向随机采集4个叶片、花和果实没有明显损伤的枝条,采集后立即放入自封袋,带回实验室测量相关指标.
-
将带回的实验材料放在恒温鼓风干燥箱中,在70 ℃下条件下烘干48 h后至恒质量,用FA1004万分之一电子天平测量茎、叶、花柄、花被、雄蕊与雌蕊群、果柄、果皮和种子干质量.生物量分配指标计算方法:用各构件的生物量占该小枝总生物量的百分比作为构件生物量分配的数量指标,即某一构件生物量分配=(该构件生物量/该小枝的总生物量)×100%.繁殖分配与营养分配指标计算方法:①花枝营养器官干质量为茎、总叶和花柄干质量之和;繁殖器官干质量为花干质量,即花柄、花被干质量和雄蕊与雌蕊群干质量之和;营养投入干质量为茎、叶、花柄和花被干质量之和;繁殖投入干质量即为雄蕊与雌蕊群干质量;②果枝营养器官干质量为茎、叶和果柄干质量之和;繁殖器官干质量为果实干质量,即果柄、果皮和种子干质量之和;营养投入干质量为茎、叶、果柄和果皮干质量之和;繁殖投入干质量即为种子干质量.
-
利用IBM SPSS Statistics 20.0软件对生物量分配参数进行方差分析,用新复极差法(D)进行多重比较.对于小枝繁殖分配与营养分配的相关性分析使用的方程y=bxa,研究前将两边取对数转化为线形方程lgy=lgb+algx,x与y分别表示小枝功能特征参数,b代表截距,a为斜率,表示异速生长方程的参数.采用标准化主轴估计(standardized major axis,SMA)方法估计异速生长方程的参数,运用软件(S)MATR Version 2.0完成,根据Pitman方法计算回归斜率的置信区间.所有绘图均在Origin 9.1软件中完成.
1.1. 研究区域
1.2. 样品采集
1.3. 指标测定
1.4. 数据分析
-
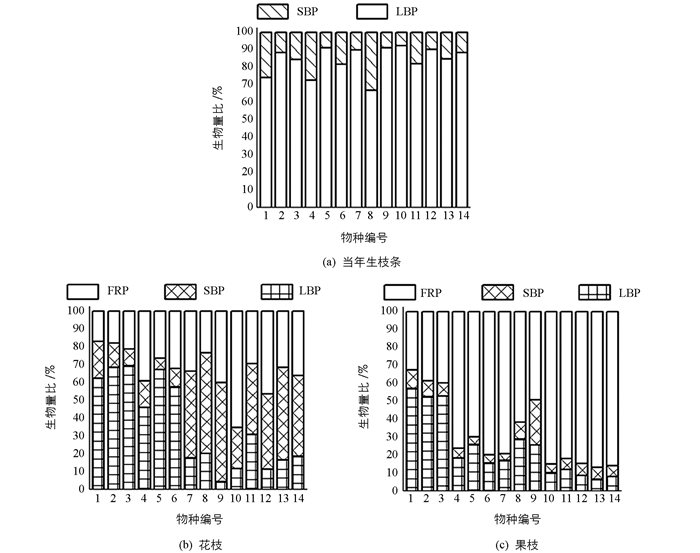
茎、叶、花和果实生物量分配差异极具有统计学意义(p<0.01).当年新生枝条叶生物量比(LBP)大于茎生物量比,其中玉兰的叶生物量比最高(即茎生物量比(SBP)最低),紫叶李相反(图 1(a));花枝中红枫、天竺桂、樟、红花檵木、海桐、海棠花分配给叶的生物量最多,李、紫叶李、紫荆、梅、紫叶桃花、桃茎生物量比最大,而玉兰、红花碧桃的繁殖器官生物量比(FRP)最大(图 1(b));果枝中除红枫、天竺桂、樟外,其余植物分配给繁殖器官、叶、茎生物量逐渐降低(图 1(c)).
-
紫荆、玉兰、梅的叶生物量比随新—花—果梯度变化先显著下降后逐渐升高,但果枝叶生物量比仍低于当年新生枝条,其余植物逐渐下降;红枫、天竺桂、樟、红花檵木、海桐和海棠花的茎生物量比逐渐下降,其余植物为先升高后下降;繁殖器官生物量比则显著升高(图 1).
-
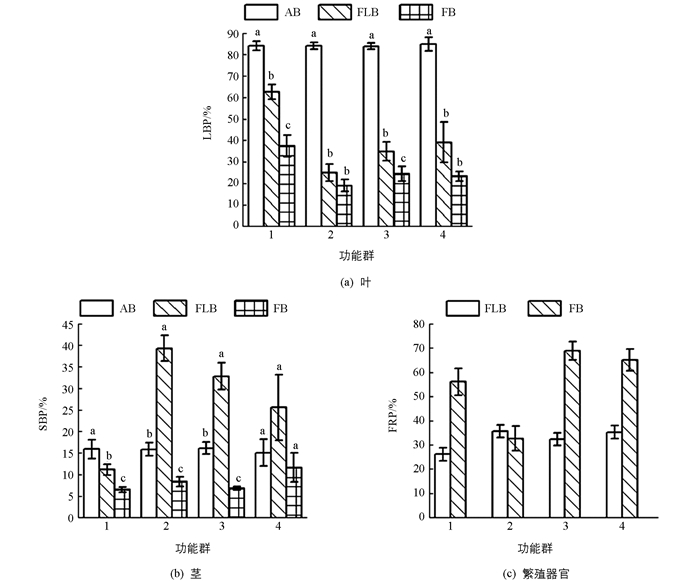
在生活型上,生物量分配在当年新生枝条上差异不具有统计学意义,在花枝及果枝中差异具有统计学意义(p<0.05).在生长型上,仅有花枝的叶生物量比和果枝的茎生物量比的差异具有统计学意义(p<0.05).当年新生枝条叶生物量分配为落叶高于常绿、灌木高于乔木,茎生物量分配相反;花枝中繁殖器官生物量分配为落叶高于常绿、灌木高于乔木,叶生物量分配为落叶低于常绿、灌木高于乔木,茎生物量分配为落叶高于常绿、灌木低于乔木;果枝中茎生物量分配为落叶高于常绿、灌木高于乔木,而叶、繁殖器官生物量分配相反(图 2).
-
除落叶植物的繁殖器官生物量比和灌木的茎生物量比外,其余生物量分配在不同发育阶段差异具有统计学意义(p<0.05).其中常绿植物、乔木和灌木的繁殖器官生物量比随发育历程的进行逐渐升高,其余生物量分配表现为下降的趋势(图 2).
-
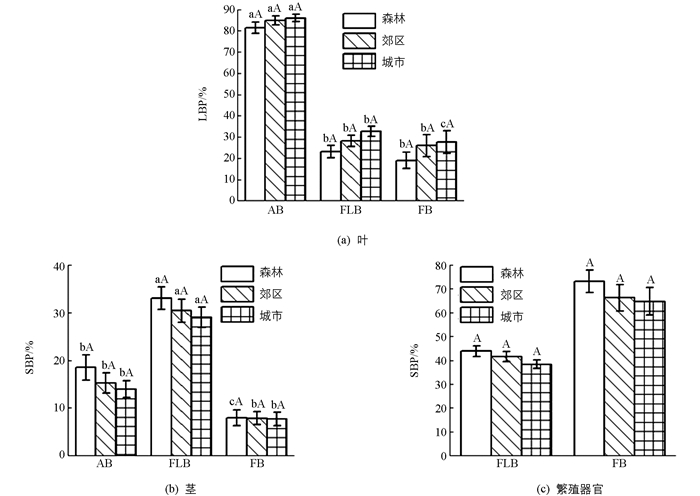
总体上,环境对生物量分配的影响不具有统计学意义.具体表现为叶生物量比随森林—郊区—城市梯度变化逐渐升高(图 3(a)),茎和繁殖器官生物量比逐渐降低(图 3(b)和图 3(c)).
-
生物量分配在不同发育阶段差异极具有统计学意义(p<0.01).具体表现为叶生物量比随新—花—果梯度变化逐渐降低(图 3(a)),茎生物量比先增加后显著降低(图 3(b)),而繁殖器官生物量比则显著增加(图 3(c)).
-
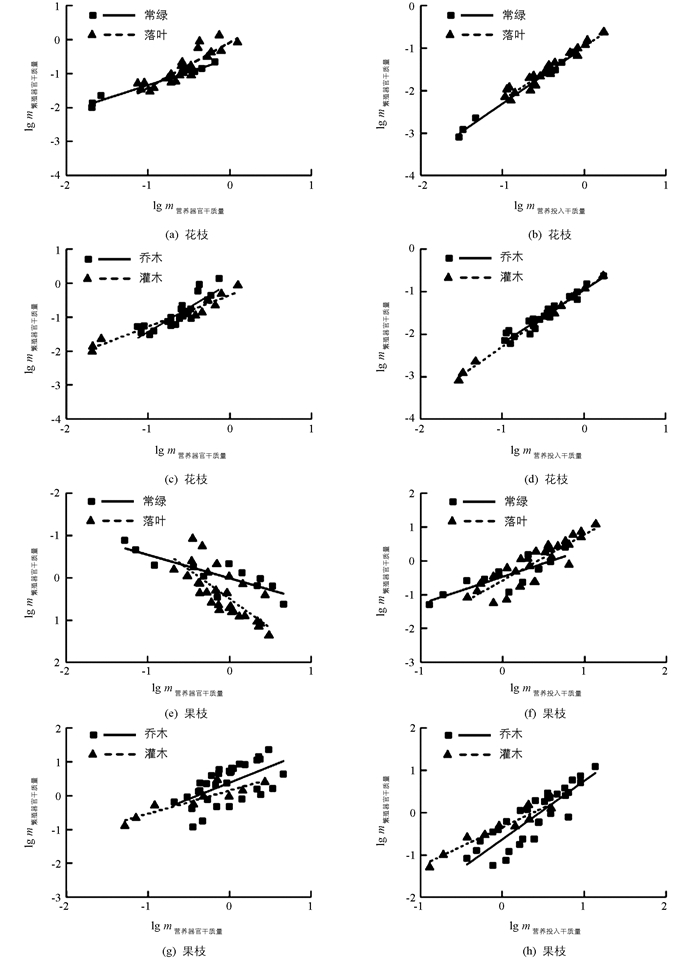
繁殖分配与营养分配在不同功能群间均呈正相关关系(表 3).在花枝中,常绿和落叶的营养器官与繁殖器官干质量斜率分别为0.788(95% CI=0.629~0.986,p=0.042)和1.487(95% CI=1.236~1.788,p<0.001)(图 4(a));营养投入与繁殖投入干质量共同斜率为1.318(95% CI=1.228~1.399,p<0.001,图 4(b)).乔木和灌木的营养器官与繁殖器官干质量斜率分别为1.636(95% CI=1.319~2.029,p<0.001)和0.948(95% CI=0.776~1.158,p=0.551)(图 4(c));营养投入与繁殖投入干质量共同斜率为1.338(95% CI=1.273~1.397,p<0.001,图 4(d)).
在果枝中,常绿和落叶植物的营养器官与繁殖器官干质量斜率分别为0.668(95% CI=0.454~0.983,p=0.042)和1.854(95% CI=1.438~2.390,p<0.001)(图 4(e));营养投入与繁殖投入干质量斜率分别为0.942(95% CI=0.653~1.359,p=0.729)和1.551(95% CI=1.285~1.872,p<0.001)(图 4(f)).乔木和灌木的营养器官与繁殖器官干质量斜率分别为1.656(95% CI=1.235~2.221,p=0.001)和0.758(95% CI=0.522~1.100,p=0.125)(图 4(g));营养投入与繁殖投入干质量斜率分别为1.595(95% CI=1.328~1.916,p<0.001)和0.952(95% CI=0.745~1.216,p=0.650)(图 4(h)).
-
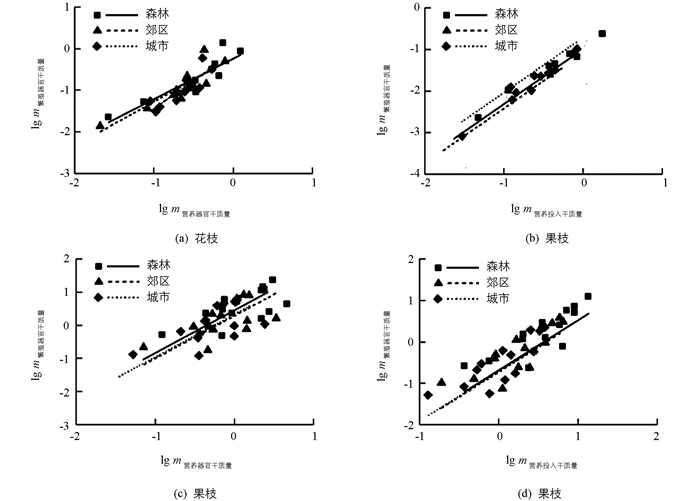
繁殖分配与营养分配在不同环境下均呈正相关关系(表 3).在花枝中,森林、郊区与城市的营养器官与繁殖器官干质量共同斜率为1.172(95% CI=0.978~1.403,p=0.084,图 5(a));营养投入与繁殖投入干质量共同斜率为1.319(95% CI=1.230~1.413,p<0.01,图 5(b)).此外,3种生境下y轴截距及沿着共同主轴的位移的差异均不具有统计学意义.
在果枝中,不同生境的营养器官与繁殖器官干质量共同斜率为1.288(95% CI=0.987~1.683,p=0.062,图 5(c));营养投入与繁殖投入共同斜率为1.204(95% CI=1.064~1.365,p=0.004,图 5(d)).此外,森林中的物种比另外2种生境的物种沿着共同主轴有显著向上的位移(p=0.017),即森林中物种具有更大的营养器官、繁殖器官、营养投入与繁殖投入干质量.
-
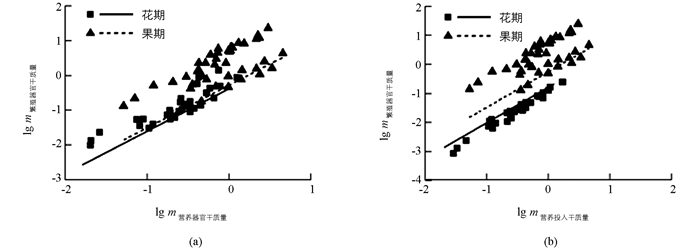
繁殖分配与营养分配在不同发育阶段均呈正相关关系(表 3).营养器官与繁殖器官干质量共同斜率为1.222(95% CI=1.074~1.395,p=0.003,图 6(a)).此外,果枝比花枝的物种沿着共同主轴有显著的向上的位移,表明果枝具有更大的营养器官和繁殖器官干质量.
营养投入与繁殖投入干质量共同斜率为1.329(95%置信区间CI=1.255~1.407,p<0.01,图 6(b)).此外,果枝的y轴截距显著大于花枝(p<0.01),且果枝沿着共同主轴有显著的向上的位移,表明果枝具有更大的营养投入与繁殖投入干质量.
2.1. 物种生物量分配总体差异特征
2.1.1. 不同物种生物量分配的差异
2.1.2. 物种在不同发育阶段生物量分配的差异
2.2. 不同功能群生物量分配的差异
2.2.1. 功能群在同一发育阶段生物量分配的差异
2.2.2. 功能群在不同发育阶段生物量分配的差异
2.3. 不同生境下生物量分配的差异
2.3.1. 不同生境下同一发育阶段生物量分配的差异
2.3.2. 不同生境下不同发育阶段生物量分配的差异
2.4. 繁殖分配与营养分配的相关性
2.4.1. 不同功能群间繁殖分配与营养分配的相关性
2.4.2. 不同生境下繁殖分配与营养分配的相关性
2.4.3. 不同发育阶段繁殖分配与营养分配的相关性
-
本研究中,不同生活型间生物量分配的差异具有统计学意义(图 2).当年新生枝条和果枝中落叶植物和灌木的叶生物量分配分别高于常绿植物和乔木,或许是前者需要较高的温度和更多光照来进行光合作用[9-10],而花枝中紫荆、桃等落叶植物在开花的同时叶片还未长出或正在萌发,可能是开花习性对生物量分配有一定的影响,在未来需进一步研究生物量分配在不同开花习性植物之间的差异.此外,花枝中落叶植物和灌木对于繁殖构件投资增大以增加繁殖成功率[11];相反,常绿植物和乔木分别通过增大叶和茎生物量分配来占据光资源优势[12],这与夏晨诚等[13]研究结果一致.果枝中落叶植物和灌木主要通过增大营养构件分配来增强竞争力,从而实现资源的最优化配置,以保证植物生长需要和提高种群繁衍的能力[14].
不同功能群间繁殖分配与营养分配的相关性差异均具有统计学意义(花枝中营养投入与繁殖投入干质量除外)(图 4).常绿植物的营养器官与繁殖器官干质量呈显著小于1.0的异速生长关系,而落叶植物则显著大于1.0,或许是在相同资源条件下,常绿物种在生长季要遭受更大的拉力,而部分落叶植物在花期叶未完全展出或果期脱落叶片[15-16].此外,乔木繁殖分配与营养分配呈显著大于1.0的异速生长关系,而灌木呈等速生长关系(花枝中营养投入与繁殖投入干质量除外),表明灌木通过增大对机械支撑或者光合、蒸腾作用的投资以占据光资源优势[17].可能是乔木和灌木由于生态位的差异,分别增大对繁殖分配和营养分配的投资,体现了同一环境中不同林层为获取更大的光资源而在繁殖分配和营养分配之间产生的一种权衡策略[17].
-
本研究中,生境对生物量分配影响无统计学意义(图 3).可能因为:第一,植物生物量分配受多种因素的影响,与植物种类、大小和温度等都有关系,植物通过调节各个器官占总生物量的分配比来适应不同的环境;第二,森林中温度较低,风力较强,植物通过增大茎和繁殖器官生物量投入来满足物种的生存和繁衍[18];第三,城市中污染高,植物通过减小繁殖器官构件分配来增强竞争力,这是植物长期面对胁迫环境而产生的一种适应性策略.
总体上,繁殖分配与营养分配相关性在不同生境中均存在共同斜率(图 5).表明在不同生境中,植物采用较为一致的繁殖分配策略,这些变化趋势与张茜等[19]的研究结果相反,可能是因为生物量分配与植株个体、污染程度、土壤性质等因子均有密切关系.此外,果枝中森林中的物种具有更大的营养与繁殖器官及营养与繁殖投入干质量.可能是森林环境中资源丰富,植物光合作用同化的产物增多,从而形成较大的叶片、花和果实[20];反之,城市环境中资源相对匮乏,植物通过减少营养和繁殖构件的储存能力,从而提高自身的存活率.
-
本研究发现,繁殖分配与营养分配在不同发育阶段具有共同斜率,且表现为呈大于1.0的异速生长关系(图 4).即繁殖构件越大的物种,在保证植物生长发育和种群繁衍上需要小枝、叶片付出更高的代价.果枝具有更大的营养分配和繁殖分配比例,其原因可能是各物种的营养分配和繁殖分配的异速生长起始点(截距)的可塑性较高,但繁殖分配在相对生长中对于发育历程变化不太敏感[21],我们认为这是物种在应对发育历程的改变而产生的一种适应性机制.此外,随着发育历程的进行,叶、茎生物量分配降低,而繁殖器官生物量分配显著增加(图 3).即在生物量分配方面,植物均采用较为一致的分配方式来适应资源变化及竞争排斥.