


 下载:
下载:
-
竞争是生态系统中生物个体间因为有限的空间或者资源而产生的相互制约的现象[1].竞争可以发生在相同物种之间(种内竞争)和不同物种之间(种间竞争)[2].种内竞争与种群密度密切相关,在植物群落中,当种群密度增大时,种内竞争强度过大可能使部分植物个体死亡,导致自疏现象[3].种间竞争主要由于一个物种的个体受到另一个物种的个体对其生长、存活等方面能力的影响.种内竞争和种间竞争共同决定着群落内种群的动态变化,影响群落内物种的组成以及结构.因此,探究物种的竞争关系,以及竞争对植物生长的影响,对于阐明物种共存以及群落物种组成和结构变化具有重要的意义.
Hegyi单木竞争模型在评估林木竞争强度方面应用广泛[4-8],张跃西等[9-10]对Hegyi竞争指数模型进行了改良,增加了竞争指数的变异程度.段仁燕等[11]通过张跃西的改进模型计算了太白红杉林的种内种间竞争强度,并结合太白红杉的胸径大小,很好地预测了随着林木生长太白红杉种群的种内种间竞争强度的变化.利用Hegyi竞争模型,巢林等[12]也成功预测了不同林龄杉木人工林林木生长过程中的竞争强度变化.
川西亚高山地区的秋季彩叶森林群落因其良好的美学景观价值和生态服务功能,已经成为了当地重要的森林旅游资源之一.不同坡向上由于群落组成结构的差异,其秋季彩叶林呈现的色彩及景观质量存在较大差异,这对当地景观林抚育维持及旅游基础设施建设造成了一定困扰.目前主流观点认为,阴阳坡上光照、水分以及温度等环境因素的差异影响了群落的结构和物种多样性,最后形成不同的景观类型[13-19].阴阳坡光照、水分以及温度等环境因素像一个“筛子”一样,决定着物种的竞争能力,影响物种间或个体间的竞争关系,进而影响森林群落的构建和森林景观的形成[20].但不同坡向上彩叶林群落内部的树木竞争模式是否存在差异尚不清楚.
为进一步阐明川西亚高山彩叶林群落在不同坡向上的植被差异和景观差异形成的原因,本文以川西地区不同坡向上彩叶林群落为研究对象,采用Hegyi竞争指数模型定量分析了阴坡和阳坡彩叶林群落内部种内和种间竞争差异以及与树木生长的关系,为彩叶林景观可持续管理与提升奠定理论基础.
全文HTML
-
研究地点位于四川省阿坝藏族羌族自治州理县沙坝村(N31.51°,E102.90°),海拔约为2 400 m.该区域是典型的高山峡谷地貌,地形以石质山地为主,坡度多在25°以上.年平均气温约12 ℃,年平均降雨量584 mm.植被以次生落叶阔叶林为主,主要优势种有华山松(Pinus armandii),青榨槭(Acer davidii),五裂槭(Acer oliverianum),橿子栎(Quercus baronii),黄栌(Cotinus coggygria),川梣(Fraxinus sargentiana)等.林下灌木主要有川桂(Cinnamomum wilsonii),黄栌(Cotinus coggygria),菝葜(Smilax china),草本层主要有苔草(Carex spp.),唐松草(Thalictrum aquilegifolium)等.
-
选取区域内人为干扰最小、位于同一海拔梯度但坡向相反的天然次生林作为研究区域设置样地.在位于不同坡向的林内分别设置3个50 m×50 m的样方,共6个.对于每个样方内总株数大于5的树种选定为研究目标物种,树木胸径≥2 cm的设定为对象木,记录对象木的胸径、树高以及冠幅,并且进行挂牌编号.以树冠接触和林木的遮荫状况来确定竞争范围,竞争范围内的个体为竞争木,实测竞争范围内对象木与竞争木之间的距离.
记录样地海拔、坡度、坡向等地形因子.于2018年8月用HH2土壤湿度计测仪测量土壤温度和土壤湿度,用Hyelec温湿计测量大气温度和大气湿度,阴坡的空气湿度和土壤湿度都要高于阳坡,其基本信息见表 1.
-
物种多样性指数选择辛普森多样性指数(Simpson Index)、香农—威纳指数(Shannon-Weiner Index)和Pielou均匀度指数.计算公式为
其中,D表示辛普森多样性指数,H表示香农—威纳指数,E表示Pielou均匀度指数,S表示样地物种数目,Pi表示种i的个体在全部个体中的比例.
利用伯杰—派克优势度指数(Berger-Parker Dominance Index),将优势指数值大于0.1的物种记为优势种,反之,为非优势种.计算公式为
其中,d为优势指数,Nmax为优势种的种群数量,NT为全部种的种群数量之和.
-
本文采用张跃西[9]改进的Hegyi单木竞争模型计算竞争指数来评估竞争强度的大小.其计算公式为
其中,CI为竞争指数,该值越大,竞争强度越大;Di为竞争木胸径;D为对象木胸径;di为对象木与竞争木之间的距离;N为竞争木的株数.先计算出每个竞争木对对象木的竞争指数,然后将N个单木间的竞争指数累加和平均即得到对象木受到的竞争强度.
当竞争木与对象木树种相同时,其竞争强度定义为种内竞争强度;当竞争木与对象木为不同树种时,其竞争强度定义为种间竞争强度.种内竞争强度和种间竞争强度之和为该对象木的竞争强度.将N株竞争木的竞争指数累加即得到该树种受到的种内竞争强度和种间竞争强度.样方内该树种的所有对象木的种内竞争强度或者种间竞争强度的值累加的平均值即得到该物种在该样地的平均种内竞争强度或者种间竞争强度.
1.1. 研究区概况
1.2. 样地设置与调查
1.3. 物种多样性指数计算以及优势物种
1.4. 竞争强度
-
首先,利用线性混合模型分别分析了竞争指数和林木个体的胸径、树高和冠幅的关系.在模型中,竞争指数为因变量,胸径、树高或冠幅为固定效应,树种作为随机效应;其次,为了综合评价林木的生长情况,将收集到的所有林木胸径、树高和冠幅进行单位Z分数标准化,三者算数平均和作为林木综合生长的指标;第三,利用单因素方差分析比较不同坡向上共同属林木生长指标的差异.
除此之外,以所有林木胸径均值为界,将胸径大于均值的林木个体划分为大径级个体,反之则为小径级.然后采用线性混合模型分析不同坡向和不同径级上所有物种林木生长与竞争强度的关系.在模型中,竞争强度为因变量,径级、坡向以及其交互项作为固定效应,物种作为随机效应. R version 3.6.1[21]用来完成本实验中所有数据分析,其中线性混合模型用lme4包来进行拟合[22],作图在EXCEL 2016中完成.
-
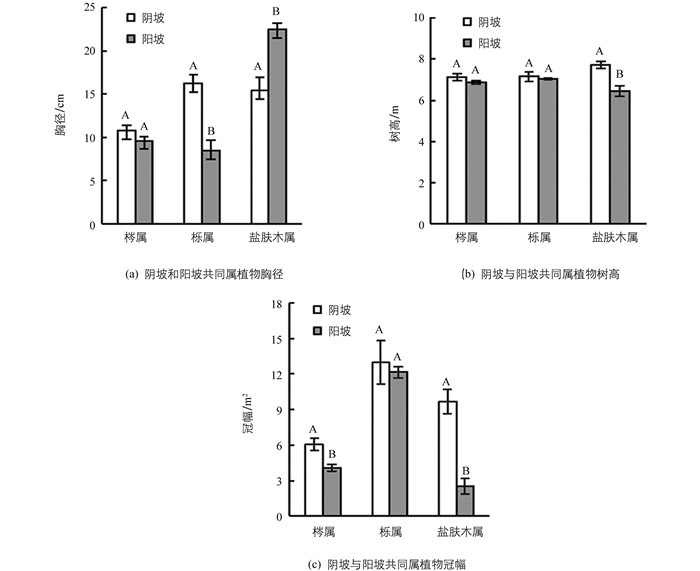
根据计算的Simpson指数和Shannon-Weiner指数,阴坡物种多样性高于阳坡.阴坡植物群落Pielou均匀度指数高于阳坡(表 2).阳坡上林分密度大约为1 146.7株/hm2,而阴坡上林分密度大约为730.2株/hm2(表 3).利用伯杰—派克优势度指数确定阴坡群落林木优势种5种,为华山松(P. armandii)、青榨槭(A. davidii)、青麸杨(Rhus potaninii)、亮叶桦(Betula luminifera)和川梣(F. sargentiana).阳坡林木优势种3种,包括橿子栎(Q. baronii)、川梣(F. sargentiana)和黄栌(Cotims coggygria).阴坡上优势物种中胸径、树高和冠幅最大的为华山松,阳坡中优势物种橿子栎胸径、树高和冠幅最大(表 3).
虽然阴坡上林木平均胸径稍低于阳坡,但是阴坡上林木平均树高和平均冠幅均大于阳坡,阴坡上林木平均综合值也大于阳坡(表 3),说明整体阴坡上林木生长优于阳坡.不同坡向上共同生长的属的植物冠幅阴坡大于阳坡,除盐肤木属植物外其他植物胸径阴坡大于阳坡,但盐肤木属树高阴坡大于阳坡(图 1).
-
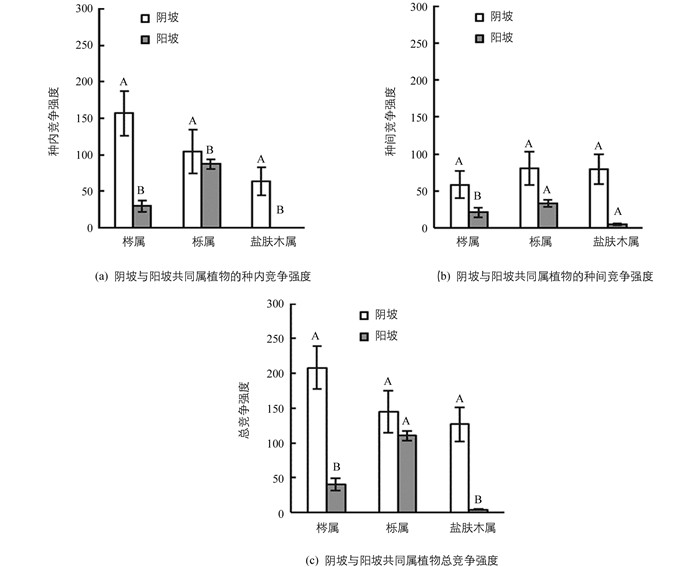
总体上看,阴坡上群落林木平均种内竞争强度、种间竞争强度都大于阳坡(表 3).比如,从阴坡到阳坡,盐肤木属、梣属和栎属植物种内、种间竞争强度表现出不同程度的下降(图 2).阴坡上优势物种竞争强度由大到小依次为川梣、亮叶桦、华山松、青麸杨、青榨槭;其中亮叶桦、川梣种内竞争大于种间竞争,青榨槭和青麸杨种间竞争大于种内竞争,华山松种内种间竞争强度差异不大.阳坡上优势物种橿子栎的林分密度是川梣的3.5倍,其竞争强度明显高于川梣,两者的种内竞争强度都大于种间竞争强度(表 3).
-
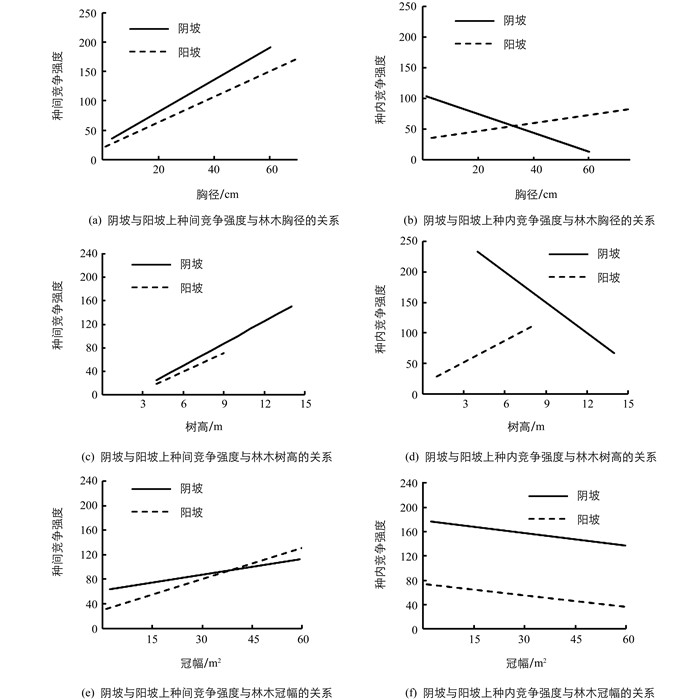
随着树木个体生长,包括胸径、树高和冠幅的增加,种间竞争强度在阴坡和阳坡上都随之增加(图 3).在阴坡上,种内竞争强度随着胸径、树高的增加而减少,但是在阳坡上种内竞争强度随着胸径、树高的增加而增加.种内竞争强度随着冠幅的增加而减少,不同坡向间差异无统计学意义(图 3).
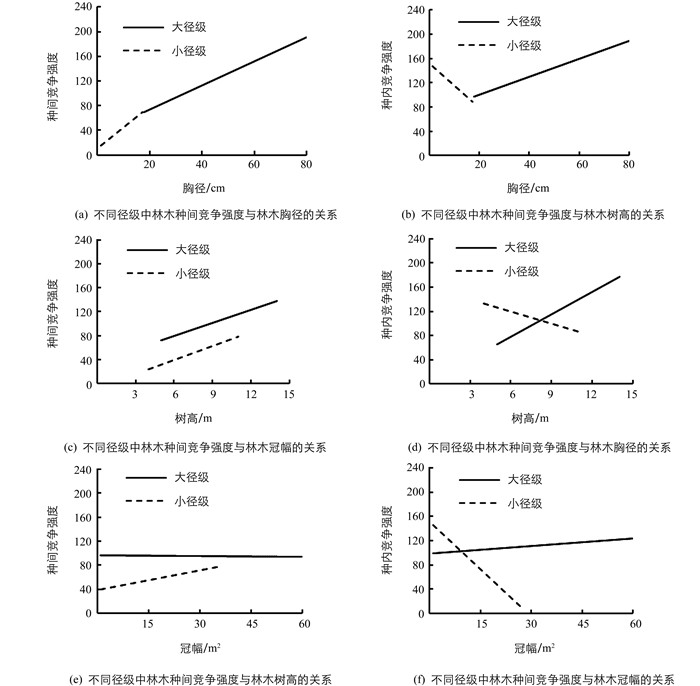
对于不同径级的林木,随着个体大小的增加,总体上种间竞争强度都呈现增加的趋势.但是,不同径级大小的林木种内竞争强度与林木生长指标的关系却显著不同(图 4).结果发现,小径级林木的种内竞争随着胸径、树高和冠幅的增加而减小,大径级林木的种内竞争强度随着林木胸径、树高和冠幅的增加而增加(图 4).
3.1. 不同坡向上群落物种多样性
3.2. 不同坡向上植物竞争差异
3.3. 不同坡向上竞争强度与林木生长的关系
-
研究发现从阴坡到阳坡,林分密度、群落组成和结构发生了明显变化.阴坡上植物群落物种多样性高,群落垂直结构层次丰富,阳坡与之相反(表 3).比较不同坡向上群落物种的种内种间竞争强度关系发现,阴坡上优势物种青榨槭、青麸杨种内竞争强度小于种间竞争强度,华山松种内种间竞争强度差异不大,但是阳坡优势物种橿子栎和川梣都表现出种内竞争强度大于种间竞争强度(表 3).相似物种可共存的前提是种内竞争大于种间竞争[23],这说明阳坡优势物种在群落中能够稳定共存,而阴坡上优势物种间关系不稳定,这可能是阴坡物种多样性高于阳坡的原因之一.
经典群落构建理论认为,环境筛选和生物间的相互作用是影响群落构建的两个主要因素[24-25].在群落演替早期环境筛选作用较强,决定群落的物种组成[26-27].在山地地形中,随着坡向的改变,太阳辐射强度、土壤理化性质、气温等生态因子等都随之改变[28].在研究区域内,阴坡的空气湿度和土壤湿度都要高于阳坡(表 1).因此坡向可能通过环境筛选作用来影响物种组成,进一步影响群落的结构和组成[29].随着群落演替进程、种群密度和物种数目增多,物种间生态位重叠,种内种间竞争强度加剧,逐渐成为演替后期的主导因素[24-25, 30-33].种内种间竞争共同影响着群落内种群的动态变化,影响群落的结构和组成[34].在研究区域内,阴坡上林木平均种内竞争强度均高于阳坡(表 3),因此阴坡上林木可能因为种内竞争强度过大出现更多生态位重叠[35],导致了自疏现象,种群密度显著低于阳坡上的种群密度.
进一步分析林木竞争强度与林木生长关系发现,阴坡上种内竞争强度随着林木胸径和树高的增加而减少,阳坡上随着林木胸径和树高的增加而增加(图 3).说明坡向显著改变了种内竞争强度与林木生长的关系,这可能是因为不同坡向之间的资源环境差异造成的.在阴坡上,大气湿度和土壤湿度都比阳坡高(表 1),生长环境优越,水分胁迫较低,林木生长可能主要受到林木的种内竞争的影响.因为种内竞争主要是由于相同物种的资源利用的普遍重叠导致的[20],随着林木不断生长,其胸径、树高和冠幅不断增加,资源利用重叠越来越大,种内竞争亦越来越激烈.在激烈的竞争环境条件下,导致了自疏现象,种群密度逐渐降低,种内竞争强度也随之降低.但是由于阳坡其生长条件较为恶劣,林木生长估计主要受到环境作用的限制,种内竞争效应较弱,引起的自疏效应不明显,因此种内竞争强度没有下降反而随着个体大小增加而增加.
另外,不同大小径级的林木其种内竞争强度与林木生长关系差异也有统计学意义.具体表现为小径级林木随着胸径、树高和冠幅的增加其种内竞争强度减少,而大径级林木随着其胸径、树高和冠幅的增加其种内竞争强度增加(图 4).有研究发现,对于同一竞争强度,大树和小树的竞争敏感性不同,对小树生长的影响更大[36].因此,随着种内竞争强度的增加,主要是小径级林木的生长受到抑制,在长期竞争环境下,种内竞争引起自疏效应主要发生在小径级林木中,而大径级林木的生长和密度没有受到影响.与种内竞争相反的是,对于不同大小径级的林木,种间竞争强度都随着个体大小的增加而增加.这说明彩叶林种群落的自疏现象可能主要是由于种内竞争引起的.
综上,在阴坡上林木生长环境优于阳坡,但是其林木平均种内和种间竞争强度均高于阳坡.在激烈的竞争环境下,尤其是种内竞争,导致了自疏现象,从而降低了阴坡上的种群密度.在阳坡上,竞争强度低,自疏现象不明显,种群密度较高,林木生长主要受到生长环境的限制.另外,研究表明当某个群落内部种间竞争大于种内竞争时,群落内部的动态变化加强,物种多样性增加[37].在阴坡上,优势物种青榨槭和青麸杨的种间竞争强度大于种内竞争强度,但是在阳坡上所有优势物种种间竞争强度都小于种内竞争强度.因此,在阳坡上群落中的相似物种或许能够更好地稳定共存,阴坡上群落内部具有较高的物种多样性.这些结果表明坡向通过影响坡上植物竞争与林木生长的关系显著改变了川西彩叶林群落的物种多样性及物种的组成和结构,进而可能影响彩叶林景观及其生态服务功能.