


 下载:
下载:
-
开放科学(资源服务)标志码(OSID):

-
农田生态系统生活着种类繁多且数量巨大的各种节肢动物,这些节肢动物提供的生态系统服务(如生物控制、授粉等)是农业可持续发展的重要条件,为人类带来巨大的经济利益[1-3]. 集约化经营的农业生产方式为保障粮食生产和供给发挥了巨大作用,却导致农田生态系统生物多样性的下降和生态系统服务功能的减退,也是传粉昆虫及传粉服务功能急剧下降的主要原因之一[4-5]. 昆虫传粉是生态系统中传粉服务功能最重要的媒介形式,也是形成和制约生态系统发挥服务功能的关键因素,传粉昆虫的种类多少和传粉质量对依赖动物传粉的作物产量和质量至关重要,传粉昆虫也为农业生态系统和自然生态系统的健康状况提供预警和反馈[6-9].
贵州省正在大力发展茶叶产业,目前全省的茶园面积已达46.67万hm2,贵州省茶叶产业将生态、有机、干净茶作为基本出发点和追求目标. 贵州处于我国生物多样性较为丰富的西南地区,山区地貌是贵州地貌的基本属性,因此贵州的大部分茶园表现为林中有茶、茶中有林的镶嵌林窗式茶园分布格局,茶园周边的自然、半自然植被在春、夏、秋3季基本上都有不同的植物次序开花,因此生活在这些自然、半自然林地的传粉昆虫种类较为丰富. 同时,由于林地和茶园的镶嵌林窗式分布格局,使得茶园成为许多传粉昆虫的重要生境,然而,在贵州开展茶园传粉昆虫的调查和研究报道相对缺乏. 因此本研究在贵州2个重要茶叶产区(都匀毛尖和湄潭翠芽)的核心地带,利用节肢动物研究常用的色盆法,对多年未使用化学农药的生态茶园开展了茶园传粉昆虫多样性调查和研究,以期初步了解贵州茶园生境传粉昆虫多样性基本情况,为贵州大力打造干净茶和生态茶提供一定的数据支撑; 同时,通过比较白、黄、蓝3种色盆对茶园传粉昆虫的诱集效果,为今后开展茶园传粉昆虫的研究选择合适的方法提供依据.
全文HTML
-
本次调查的2个生态茶园分别属于都匀毛尖(黔南州龙里县)和湄潭翠芽(遵义地区湄潭县)的主产区. 龙里茶园位于东经107°2′8″,北纬26°13′29″,海拔1 277 m; 湄潭茶园位于东经107°32′19″,北纬27°39′48″,海拔约886 m. 2个茶园均位于山区,与其周围的树林构成了镶嵌分布的林窗式生态茶园. 总体而言,龙里茶园周边的自然、半自然生境面积明显多于湄潭茶园,且龙里茶园地貌多样性优于湄潭茶园.
-
调查采用色盆诱集法、又称彩盆法或挂盆陷阱法[10],色盆为市售一次性圆形透明塑料快餐盒用自喷漆自制而成. 色盆上口内径13 cm,下口内径8.5 cm,高8 cm,自喷漆颜色分为珍珠白(pearl white)、美术黄(canary yellow)、天蓝(sky blue)3种颜色,将塑料盒的内外均匀喷好漆后晾干备用. 使用时盆内加入400 mL水、1.2 g苯甲酸钠防腐剂及1滴液体清洁剂,色盆放置在茶蓬冠层上,并保持色盆上口与冠层上部齐平. 在一条茶蓬上3种颜色的色盆各放一个,色盆间距在3 m左右. 试验设3次重复,即3条茶蓬,重复间相隔5条茶蓬.
色盆在茶园中放置7 d后收回,调查频率为每月1次,从5月至10月共调查6次.
-
本文所述的传粉昆虫包括:膜翅目、双翅目(食虫虻总科、食蚜蝇总科和蝇总科)、鞘翅目、半翅目(显角亚目)、缨翅目、鳞翅目昆虫[11-13].
所收集标本均带回实验室进行分类鉴定和统计分析. 数据整理在Excel上进行,物种丰富度、群落多样性(Shannon-Wiener index)香农指数、均匀度指数和群落相异系数均在DPS 18.10软件上进行统计分析.
Shannon-Wiener多样性指数(H′):
Pielou均匀度指数(E),基于Shannon-Wiener多样性指数计算,其中Hmax=lbS.
群落相异系数即Jaccard指数(Cj):
式中,Pi为第i个物种个体数量占群落中总个体数量的比例,Ni为第i个物种的数量,N为群落个体总数; lb为以2为底的对数计算,S为群落的物种总数; a,b为两群落各自的物种丰富度,j为两群落共有物种丰富度. 根据Jaccard指数的原理,当Cj为0~0.25时,两群落为极不相似,Cj在0.25~0.50时,为中等不相似,Cj为0.50~0.75时,为中等相似,Cj在0.75~1.00时为极相似.
1.1. 调查地点概况
1.2. 调查方法
1.3. 数据处理与分析
-
总体而言,龙里茶园诱集到的物种丰富度和个体数量均高于湄潭茶园. 在龙里茶园共采集到2 225头传粉昆虫,分别属于58科、119种. 其中白盆诱集到的传粉昆虫物种丰富度和香农指数均为最高,分别是73种和2.68; 蓝盆诱集的物种丰富度最低,仅为47种,而数量最高,为854头,相对而言,其香农指数和均匀度均为最低. 在湄潭茶园采集到1 658头传粉昆虫,分别属于41科、76种. 其中白盆和黄盆诱集到的物种丰富度和个体数量基本一样,白盆和黄盆物种丰富度分别为44种和45种,个体数量分别为710头和767头; 蓝盆只诱集到35种,个体数量也明显低于白盆和黄盆,诱集数量仅为181头,但是蓝盆的香农指数和均匀度指数均高于白盆和黄盆(表 1).
-
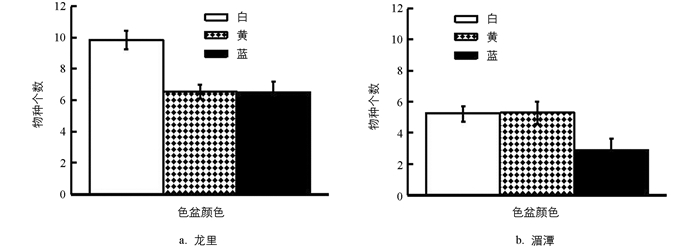
试验分析了龙里和湄潭每个色盆每次取样时诱集到的传粉昆虫物种丰富度. 结果看出,龙里茶园白盆诱集到的物种丰富度高于黄盆和蓝盆,且差异有统计学意义(p=0.0096),白盆平均每次诱集到接近10种传粉昆虫,黄盆和蓝盆诱集到的物种丰富度几乎一样; 而湄潭茶园白盆和黄盆诱集到的物种丰富度相似,蓝盆每次诱集到的物种丰富度仅为3种,但是3种色盆之间的物种丰富度差异无统计学意义(p=0.0564)(图 1).
-
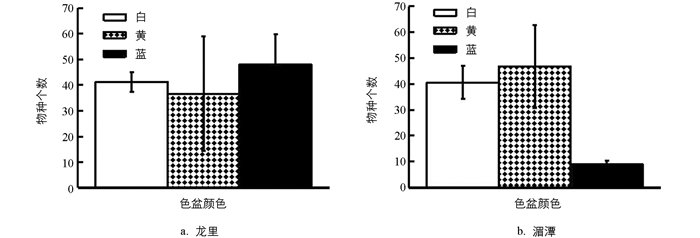
试验统计了3种色盆在龙里和湄潭茶园每次取样时的诱集数量,结果可以看出,龙里茶园的3种色盆诱集数量差异无统计学意义(p=0.549 4),而湄潭茶园的蓝盆诱集数量低于白盆和黄盆的,且差异有统计学意义(p=0.022 7)(图 2),主要原因是白盆诱集到了数量很高的姬小蜂,黄盆诱集到了数量很高的广腹细蜂,而蓝盆以上2种蜂均很少.
-
试验结果看出,3种色盆诱集到的种类均属于中等不相似的范围之内,说明了在2个茶园3种色盆诱集到的种类差异较大. 在龙里茶园,黄盆与蓝盆的诱集种类区别最大,相异性系数为0.261 4,基本上接近极不相似范围(0.25); 而湄潭茶园白盆与黄盆的诱集种类区别最大,相异性系数仅为0.263 9,也基本上接近极不相似范围. 说明在不同地点不同色盆诱集到的种类差别较大(表 2).
-
试验结果看出,龙里茶园中3种色盆诱集到的传粉昆虫均以膜翅目昆虫多样性最高,白、黄、蓝盆诱集到的膜翅目昆虫物种丰富度分别为27种、33种和24种,分别占其总物种丰富度的36.99%,61.11%和51.06%; 而诱集到的个体数量比例相对较低,其中白、黄、蓝盆诱集数量占总诱集数量的百分比分别为15.76%,18.56%,10.89%. 鞘翅目昆虫的物种丰富度在龙里茶园为次高类群,其中白盆诱集到了22种,远远高于黄盆的12种和蓝盆的6种; 同时,鞘翅目昆虫诱集数量的差异也较为明显,主要原因是在8月时,白盆和蓝盆的无斑弧丽金龟诱集数量分别为40.3头/盆和35.3头/盆,而黄盆没有诱集到. 双翅目昆虫的物种丰富度诱集情况与鞘翅目较为相似,也表现为白盆和蓝盆诱集到的物种丰富度高于黄盆,但是数量差异没有鞘翅目明显. 缨翅目昆虫数量是龙里茶园的数量优势类群,白、黄、蓝盆的诱集数量分别为总诱集数的36.54%,67.58%,61.48%. 主要种类为茶园常见的害虫茶棍蓟马和茶黄蓟马,2种蓟马的发生期均以5月为主,6-10月取样时,每盆诱集到的数量基本低于10头(表 3).
-
试验结果看出,湄潭茶园中3种色盆诱集到的传粉昆虫也以膜翅目昆虫类群为主,白、黄、蓝盆诱集到的膜翅目昆虫物种丰富度分别为26种、32种和24种,分别占总诱集物种丰富度的59.09%,71.11%和68.57%; 其中白盆和黄盆诱集数量的百分比也较高,分别为总诱集数量的49.44%和61.54%,而蓝盆为24.73%; 白盆诱集数量较高的物种为姬小蜂,黄盆诱集数量较多的为广腹细蜂,蓝盆则没有诱集到数量明显较多的类群. 物种多样性的次高类群为双翅目,白、黄、蓝盆诱集到的物种丰富度分别为7,5,4种,诱集到的个体数量均较少. 湄潭茶园中的鞘翅目昆虫物种丰富度和个体数量均较低. 缨翅目昆虫数量也是湄潭是茶园的优势类群,白、黄、蓝盆的诱集数量分别为总诱集数量的43.52%、32.72%和63.19%. 湄潭茶园诱集到的主要种类为茶园常见的茶棍蓟马和茶黄蓟马,数量发生高峰期在10月(表 4).
总体而言,2个茶园物种多样性最多的类群均为膜翅目昆虫,龙里茶园的物种多样性要高于湄潭茶园,主要原因是龙里茶园诱集到的鞘翅目昆虫物种丰富度和个体数量均高于湄潭茶园.
2.1. 龙里、湄潭两地茶园传粉昆虫情况
2.2. 3种色盆诱集的传粉昆虫物种丰富度比较
2.3. 3种色盆诱集的传粉昆虫个体数量比较
2.4. 3种色盆诱集的物种相异性比较
2.5. 龙里茶园不同目物种丰富度及其数量比较
2.6. 湄潭茶园不同目物种丰富度及其数量比较
-
本研究调查的2个生态茶园传粉昆虫群落总体特征为物种丰富度较高,且群落均匀度较好,在龙里茶园共诱集到119种传粉昆虫,共2 225头. 在湄潭茶园共诱集到76种,共1 658头,其中881头为膜翅目昆虫,超过了整个群落个体数量的一半. 其中白盆在湄潭茶园中诱集到了数量较大的姬小蜂(226头),黄盆诱集到了403头广腹细蜂,由于这2类昆虫不取食茶树,而且在调查中也未在这茶园中发现其潜在寄主大发生的迹象,因此推断茶园周边的生境中有其寄主大量发生,且这些寄主有较高的寄生率. 较高的物种丰富度和较为均匀的数量不仅说明了这2个生态茶园均有较为合理的群落结构,也说明了林窗式茶园与其周边自然、半自然生境有着较为频繁的物种交流和互动.
相对于湄潭茶园,龙里茶园诱集到较多的鞘翅目物种和个体数量. 尤其是白盆和蓝盆诱集到数量较大的丽金龟科无斑弧丽金龟,说明这2种颜色对无斑弧丽金龟有较好的诱集作用,今后在针对弧丽金龟的绿色防控工作中可开展相关研究工作. 无斑弧丽金龟是优质阔叶用材和高效生物防火树种木荷的主要传粉昆虫,其携粉量甚至高于中华蜜蜂[14],但目前我们只关注到了无斑弧丽金龟的负作用[15-16]. 在茶园生境中,尽管无斑弧丽金龟也可能取食茶叶,但是在茶叶生产中未见造成经济损失的相关报道,考虑到无斑弧丽金龟的传粉功能,在制定防控措施的时候也应加以考虑. 在茶园生境中诱集到大量的无斑弧丽金龟也说明了茶园生境与其周边生境有着强度较高的种群数量流动.
鳞翅目是一类十分重要的传粉昆虫,Ollerton[17]估计鳞翅目昆虫占所有传粉者物种种数的40%,而本研究获得的鳞翅目类群无论物种多样性和个体数量均很少,与鳞翅目种类是重要传粉昆虫的统计结果不太一致,我们认为导致这种差异的主要原因可能是色盆法对鳞翅目昆虫诱集效果较差. 盘诱法对蜂类而言是最为通用、适合的方法,此方法不仅采样效率高,而且不受采集者的经验影响[18-19],段美春等[10]的研究也证明了色盆法对蜂类研究好于扫网法、目测法、地陷法. 今后的类似研究工作可以考虑采用马氏网法和扫网法,这2种方法均简单且高效,可获得更加全面的传粉昆虫多样性研究结果.
3种色盆诱集到的传粉昆虫物种构成相异性较大,而且在龙里和湄潭2个地方导致相异性较大的具体类群均有所不同,在龙里茶园是鞘翅目类群的差异较大,而在湄潭茶园是膜翅目类群的差异较大. 因此在今后的研究中可以根据研究重点而使用不同颜色的色盆,例如,如果重点考虑膜翅目昆虫,则采用黄色色盆,如果重点考虑鞘翅目可以考虑白色或者蓝色色盆.
-
茶树种植的目的不是收获其开花后的果实,从而导致我们对茶园生境中活动的传粉昆虫调查和研究很少,在多山地貌的贵州,茶园基本上建在山坡地,因而茶园与其周边的自然、半自然生境呈现出镶嵌式分布格局,茶园必然是其周边各种生境的重要组成部分,因此可以预见贵州茶园与其周边的各种生境存在着较为频繁的传粉昆虫交流和互动. 正如李良涛等[20]指出,农田与半自然生境的互补是促使传粉昆虫在破碎化景观中得以续存的一种机制; 同时,农业景观中自然、半自然生境的存在能有效提高传粉昆虫的多样性与丰富度,从而提升作物的产量,有研究证明了生境复杂度更高的生境比生境简单的地区有更高的传粉昆虫多样性、丰富度和均匀度,也指出自然、半自然生境的存在能更好地维持传粉者的多样性和群落数量的稳定性[21-22]. 因此,在贵州进行茶园园区规划和建设时也应该考虑如何有效利用当地自然、半自然生境中提供的各种生态服务功能,包括生产、环境保护、野生动植物保护和景观美学功能.
传粉者物种丰富度和数量的多寡直接反映出该生态系统的健康程度,本次调查的龙里茶园采集到58科、119种传粉昆虫,而湄潭茶园仅采集到41科、76种. 本研究认为两地茶园物种多样性的差异反应出两地茶园周边生态系统的不同,例如龙里茶区的自然森林面积高于湄潭茶区,山地相对垂直差异大于湄潭茶区,村庄密度小于湄潭茶区,但是当地目前的茶园传粉昆虫数据很难将这两者的定量关系直接关联,传粉昆虫在贵州山区茶园与其周边茶园镶嵌式分布的自然、半自然生境的互动机制和互动强度值得进一步深入开展量化研究. 就目前来说,开展茶区传粉昆虫长期监测较为合适,开展传粉昆虫长期系统地监测不仅能对整个茶园所处的地区生态系统进行有效监测和预警[23],也能为今后开展量化研究和茶园生态工程规划建设提供必要基础信息. 近年来,建设农田生态系统的生态工程研究引起了越来越多的关注[24-27]. 同时,如果把生态茶园建设视为生态保护计划的一个组成部分,可有效促进经济效益、社会效益和生态效益的相互协调发展.