


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
在水华暴发的水体中,除氮、磷营养的积累外,丰富的有机质也在水环境中逐渐富集。作为常用碳源,葡萄糖几乎可以被所有生物利用。在水华暴发期间,藻源有机质(Algal organic matter,AOM)因大量浮游藻类在其生命周期内代谢渗出或细胞自溶裂解而产生,大量存在于水体中,使得水体溶解性有机质含量升高[1-2],其中胞内有机质(Intracellular dissolved organic matter,I-DOM)和胞外有机质(Extracellular dissolved organic matter,E-DOM)中80%~90%的组分是葡萄糖等碳水化合物[3-4]。文献[5]测定了韩国南部沿海的溶解性有机营养,发现甲藻水华暴发后其含量升高了38.38%。此外,文献[6]调查了2018年中国巢湖藻华前后的水溶性有机质质量比,发现该片水域沉积物的水溶性有机质质量比因藻类的大量繁殖而得到显著提高,峰值由233.59 mg/kg上升到306.64 mg/kg,增长了31.27%。文献[7]的研究表明,藻类衰亡初期会释放大量有机质,溶氧量急剧下降。因此,在水华暴发期间,藻类将长期处于有机质富裕的水环境中。
蓝细菌水华是全球面临的最主要的淡水生态问题[8]。作为一种光合自养微生物,蓝细菌通过光合作用获得细胞所需的碳水化合物[9]。作为水生环境中内源溶解性有机质的重要贡献者[10-11],部分蓝细菌也能利用环境现成有机质进行代谢产能。例如,微藻(Microalgae)在黑暗条件下可以利用现成的有机质积累生物量[12-13]。蓝细菌水华暴发期间,总有机碳(Total organic carbon,TOC)的大幅度增加会导致藻类消耗更多的O2,从而为水中磷酸盐的有效释放提供厌氧环境[14],磷营养盐的增加会更有利于藻类的生长[15]。但目前,随着富营养水体有机质含量的提高,蓝细菌细胞微形态及代谢转录水平的变化尚不明晰,蓝细菌对有机质富裕水环境的响应机制也鲜有研究报道,这影响了当前对广泛存在的富营养化水体生态环境的理解和预测。
位于内蒙古高原中西部干旱半干旱区的湖泊岱海,近年来湖泊水位逐年下降,水面积不断减少,其富营养化程度正在日益加剧,上覆水中溶解性有机碳的浓度达到了7.1×103 μmol/L[16-17](相当于质量浓度1.3 g/L的葡萄糖溶液)。据此,本试验采用BG11-葡萄糖营养液模拟富营养化水体水质[18],相应设定6个葡萄糖质量浓度梯度组,对蓝细菌集胞藻PCC 6803的生长代谢以及转录组学水平展开研究,目的在于探究富有机质水体对蓝细菌生长适应机制的实际影响,揭示蓝细菌在实际环境中的响应规律,同时进一步分析浮游植物群落演替的驱动因子。
全文HTML
-
本试验用到的集胞藻PCC 6803是一种单细胞蓝细菌,曾在古巴东部的水库和夏季营养过剩的湖泊等区域参与过淡水水华[19-22],能进行放氧型光合作用,具有天然的DNA转化系统,全基因组序列测定于1996年完成,能够广泛用于光合作用和蓝细菌生物学研究[23]。藻种由西南大学资源环境学院生物质能源转化实验室提供,藻种的转移均在无菌条件下进行。藻种基础培养基选用BG11[24]。配制溶液所用药品均为分析纯试剂。
仪器设备:KRQ-300P型人工气候箱、LDZX-40I型ALP高压蒸汽灭菌器、A75-H20型循环水真空泵、TU-1901型双光束紫外可见分光光度计(普析)、DR-8D型三孔电热恒温水槽、F310型浮游生物计数仪、DHG-9240A型电热恒温鼓风干燥箱、pHS-3C数显酸度计、JJ-CJ型超净工作台、扫描电镜17A00403(荷兰,Phenom World)。
-
PCC 6803的60 mL液体培养液BG11其组分如表 1所示。接种前的培养基均经过121 ℃、0.1 MPa高压灭菌30 min。
将培养至对数生长期的PCC 6803作为待接藻种,接种量为1%(V/V)。培养条件为(30±1) ℃,光照强度为2 000 lx,光暗周期设定12 hL-12 hD,每天早、中、晚定时摇瓶各1次。
以BG11作为基础培养基,并添加葡萄糖作为有机碳源。设置6个葡萄糖质量浓度梯度的处理组,分别为:0 g/L(CK)、0.7 g/L(T1)、1.4 g/L(T2)、2.1 g/L(T3)、2.8 g/L(T4)和3.5 g/L(T5),每个处理组设置3次重复。从第0 d开始,每隔2 d测定一次相关指标,包括藻细胞密度等,培养结束时分别收集样品进行细胞形态和转录组分析。
-
从第0 d开始,每隔2 d进行测定。测定前将培养液摇匀,取0.2 mL定时培养的藻液以适当比例稀释后,用血球计数板进行显微计数。根据公式(1)计算藻细胞密度(个/L):
式中:D代表藻细胞密度(个/L);N代表实际显微计数的总数(个);M代表物镜倍数;W代表浮游计数框宽度(cm);Vd代表稀释体积(mL);R代表实际计数的行数;De代表目镜直径(cm);Vc代表计数时吸取的体积(mL);n代表稀释倍数。
-
采用pHS-3C数显酸度计直接测定培养液的pH值。
-
采用热乙醇-比色法[25]测定叶绿素a的质量浓度。取测定藻细胞密度之后的剩余藻液,通过微孔滤膜(水系,孔径0.45 μm)真空过滤,将过滤完成的滤膜装入10 mL离心管中用热乙醇法提取色素,于4 ℃冰箱中避光6 h后,以3 500 r/min离心10 min,取上清液,适当稀释后测定,按公式(2)计算提取液中叶绿素a的质量浓度(mg/L)[26]:
式中:C1代表叶绿素a的质量浓度(mg/L);A665、A649分别为藻在波长665 nm、649 nm的吸光度。
-
浮游植物在水体中分泌碱性磷酸酶克服可利用磷的限制[27],同时碱性磷酸酶活性的增强表明藻类逐渐受到磷的胁迫作用[28]。本研究选用碱性磷酸酶活性指标来反映外界环境变化对蓝细菌集胞藻的影响。测定方法选用对硝基苯酚磷酸二钠(PNP-P)比色法,以对硝基苯酚的生成量表征酶的活性,计算公式如下[29]:
式中:a代表碱性磷酸酶的活性[μmol(h·mL)-1];C2代表生成的对硝基苯酚的量(μmol);t代表反应时间(h);V代表吸取反应溶液的体积(mL)。
-
使用扫描电镜17A00403(荷兰,Phenom World) 观察藻细胞的显微结构。预先使用实验室光学显微镜观察不同葡萄糖质量浓度下的藻细胞状态差异,再结合藻液颜色,以及考虑试验成本,选择葡萄糖质量浓度组CK、T1、T3和T5进行扫描电镜观察。
-
选取9份代表性样品(CK_1、CK_2、CK_3、T1_1、T1_2、T1_3、T3_1、T3_2、T3_3,尾数1、2、3分别表示同一处理的3个重复)分别低速离心10 min(4 ℃,3 500 r/min),离心结束后去上清,取沉淀,即PCC 6803细胞聚集物。将9份聚集物样本分别用无RNA酶去离子水清洗3遍后收集至冻存管,随后立即浸入液氮速冻,送至上海美吉生物医药科技有限公司进行转录组高通量测序工作,测序结果通过美吉生物云流程分析平台(
https://analysis.majorbio.com/ )进行分析。收样过程中所使用的器材全部提前进行去RNA酶处理。 -
提取mRNA前,首先检验藻液纯度,比对参考基因组特定序列(GCA_018845095.1;GCF_018845095.1),菌鉴报告显示CK、T1、T3组3个重复的比对结果均为Synechocystis sp. YACCYB藻种,满足上机的纯度要求。藻液纯度达标的样品被随即用mRNA富集法处理,用打断缓冲液把获得的mRNA片段化,在逆转录酶的作用下,利用随机引物,以mRNA为模板反转录合成一链cDNA,结合二链cDNA形成双链DNA,双链DNA经过尿嘧啶-N-糖基化酶(UNG酶)消化第二链,处理过后进行PCR扩增,热变性成单链直至得到单链环状DNA文库,将检测合格的文库进行Illumina测序。
-
原始测序数据首先通过SeqPrep 2011和Sickle 1.33软件去除接头及低质量的序列,之后利用RSeQC 2.3.6将获得的干净读取数据比对到集胞藻PCC 6803参考基因组,使用Bowtie 2将干净数据比对到基因组序列上,然后使用RSEM计算各个样品的基因表达水平。通过DESeq2软件进行样品组间差异表达分析,将满足阈值|log2FC|≥1且p.adjust<0.05的基因定义为差异表达基因(Differential expressed genes,DEGs),式中:FC为基因表达水平的差异倍数,并对差异表达基因进行基因本体(Gene Ontology,GO)富集分析和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析。
-
采用Origin 2021对藻类的各项生理指标作图,所有数据运用SPSS 27.0软件进行方差分析。通过单因素方差分析,并在p=0.05显著性水平上使用最小显著性差异检验来区分平均值之间的差异。
1.1. 藻种、试剂与仪器
1.2. 集胞藻PCC 6803的培养
1.3. 藻细胞密度的测定
1.4. 培养液pH值的测定
1.5. 叶绿素a的测定
1.6. 碱性磷酸酶(Alkaline phosphatase,AKP)活性的测定
1.7. 藻细胞形态
1.8. 转录组测序
1.8.1. 转录组实验样品采集
1.8.2. mRNA的提取及转录组高通量测序
1.8.3. RNA-seq分析
1.9. 数据处理
-
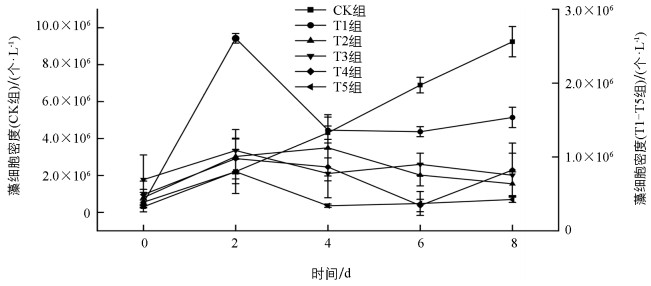
CK处理组PCC 6803的细胞生长速度明显优于其他处理组下的生长速率(图 1)。CK组的藻细胞密度增长最快,由初始值0.30×106个/L上升到8 d的9.24×106个/L,增长了约29倍;其次密度增长较快的是T1组,从初始值0.38×106个/L到8 d的1.54×106个/L,增长了约3倍。相比之下,T2-T5组的藻细胞生长速度平缓,在密度范围0~2.0×106个/L内上下波动,其中,最高峰出现在T2组的4 d,达到了1.12×106个/L,较培养起始(0 d)增长了约1.5倍。由此可知,PCC 6803的增殖生长明显受到外源葡萄糖的影响,并且当葡萄糖质量浓度超过1.4 g/L后,细胞数量生长速度明显变慢。
-
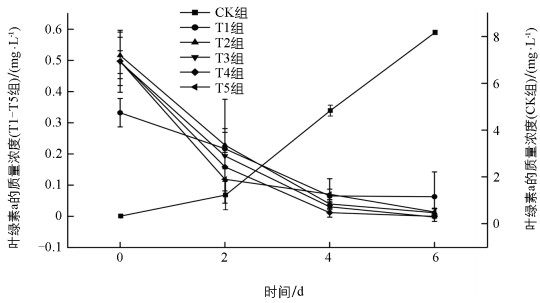
有、无葡萄糖的处理使得叶绿素a的质量浓度变化呈现相反的趋势(图 2)。CK组经过6 d培养,叶绿素a的质量浓度大幅度增长;而其他组的集胞藻叶绿素a的合成减弱,培养后期叶绿素a的质量浓度一直处于较低的水平,第6 d几乎检测不到叶绿素a的质量浓度,藻液失绿。
-
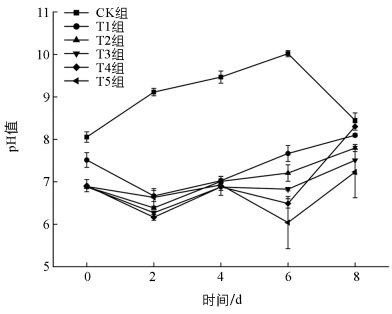
CK组的pH值在0~6 d持续增长,且均大于8.00,最大值达到了10.02;其他组的pH值均在0~2 d逐步下降,后又逐渐上升,取值范围为6.04~8.30(图 3)。
-
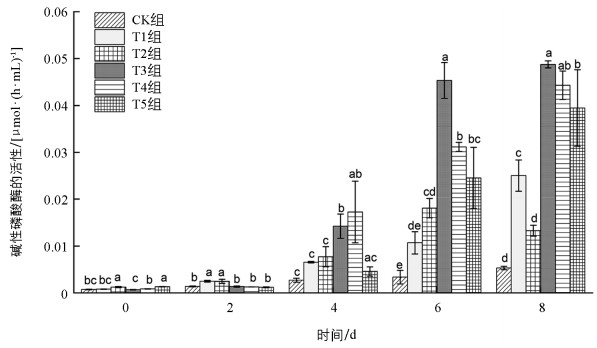
碱性磷酸酶的活性总体表现为添加葡萄糖的处理组高于CK组(图 4),并且不同处理组间的差异有统计学意义。在BG11-葡萄糖培养基中,随着培养时间的增加,处理组碱性磷酸酶的活性显著增强(p<0.05),8 d碱性磷酸酶活性由大到小的处理组依次为:T3、T4、T5、T1、T2,其数值从大到小分别为CK组的9.19、8.35、7.43、4.72及2.50倍。
-
图 5为不同葡萄糖质量浓度下(CK组、T1组、T3组、T5组)集胞藻PCC 6803放大30 000倍的扫描电镜图。从细胞大小上看,CK组、T1组和T3组的细胞大小无明显差别,T5组发生变形的细胞缩小,未变形的细胞均大于其他组别;从细胞形状上看,CK组的藻细胞呈现圆形、光滑及完整的细胞形态,细胞间无明显的丝状胞外多糖(Exopoly Saccharides,EPS),而在葡萄糖处理下,藻细胞形态开始变得不规则,T1组和T5组的藻细胞显示变形并有凹陷,T3组的细胞表面出现褶皱裂纹,并且胞外多糖显示增多。
-
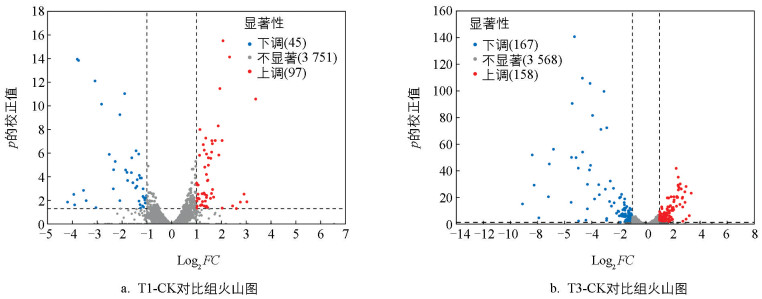
为了获得集胞藻PCC 6803于不同葡萄糖质量浓度培养组的差异表达基因,分别对CK组、T1组和T3组的蓝细菌基因进行了转录组学测序。对不同处理组的差异表达基因进行统计,发现T1组包含97个上调DEGs、45个下调DEGs;T3组有158个上调DEGs、167个下调DEGs。
针对2个对比组,进行表达水平的差异倍数(Fold Change,FC)分析,并制作火山图(图 6)。图中,蓝色和红色点分别代表表达水平显著下调和上调的差异表达基因,而灰色点表示无显著变化的差异表达基因。T1-CK和T3-CK对比组的log2FC值范围分别为-4.19~6.54和-9.11~7.20。对两个对比组进行差异表达基因水平分析,可以发现,T3组上调和下调的差异表达基因水平分别较T1组增加了62.89%和271.11%,并且部分差异基因显著性更强。
-
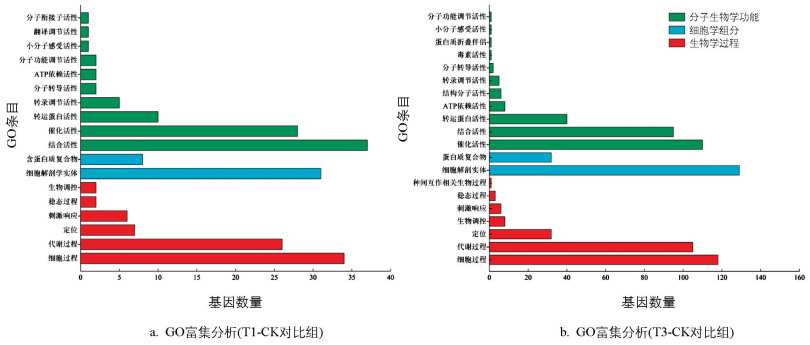
如图 7所示,在葡萄糖-蓝细菌共培养8 d后,对2个对比组的差异基因进行GO富集分析,呈现出了T1和T3处理组中丰度前18条和20条通路。T1-CK对比组涵盖了10个分子生物学功能项、6个生物学过程项、2个细胞学组分项;T3-CK对比组涵盖11个分子生物学功能项、7个生物学过程项、2个细胞学组分项。
T1组集胞藻PCC 6803的分子生物学功能呈现出显著的差异表达,其中涉及结合活性的差异表达基因有36个,涉及催化活性的差异表达基因有29个;在生物学过程中,关键的差异表达基因集中在细胞过程(34个基因)和代谢过程(27个基因);从细胞学组分方面看,差异表达基因处在细胞解剖实体(31个基因)以及蛋白质复合物(9个基因)内。与此同时,T3组在分子生物学功能、细胞学组分和生物学过程方面均与T1组呈现一致的趋势。差异表达基因在生物学过程中的细胞过程(119个基因)和代谢过程(105个基因)占比最多;其次,在分子生物学功能方面,差异表达基因主要分布在催化活性(112个基因)和结合活性(96个基因);细胞学组分结果显示,细胞解剖实体(129个基因)以及蛋白质复合物(33个基因)为差异表达基因所处的主要部位。
上述结果表明,在对葡萄糖-蓝细菌培养体系的响应中,集胞藻细胞通过调控涉及结合和催化活性功能的差异基因表达,协同调节着细胞有机体和蛋白质复合物内的基因表达功能以及蛋白质结构,进而影响自身细胞的正常生长和代谢进程。
-
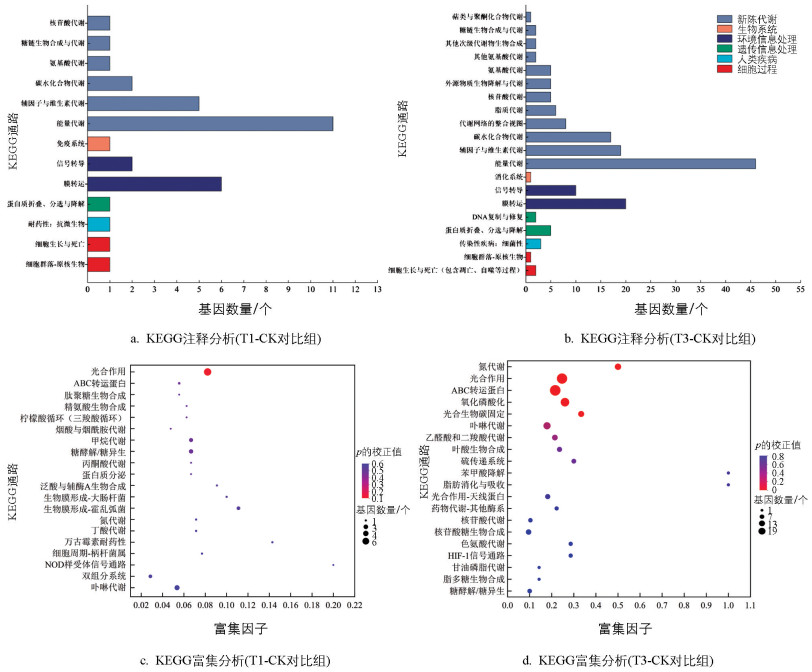
KEGG数据库致力于为生物学研究提供基因、蛋白质和代谢通路等综合信息,有助于理解基因与生物生命活动过程之间的关系。对CK组、T1组、T3组培养8 d体系的转录组进行KEGG富集分析,结果如图 8所示。图 8a、b分别展示了T1和T3组别中的蓝细菌前13条和20条通路。这些通路涵盖了生物系统、环境信息处理、遗传信息处理、细胞过程和新陈代谢等关键生物学信息。T1组蓝细菌的差异表达基因主要被注释到新陈代谢中的能量代谢(11个基因)、辅因子和维生素代谢(5个基因)、碳水化合物代谢(2个基因),以及环境信息处理中的膜转运(6个基因)和信号传导(2个基因)。T3组蓝细菌的差异表达基因主要被注释到新陈代谢中的能量代谢(46个基因)、辅因子和维生素代谢(19个基因)、碳水化合物代谢(16个基因),以及环境信息处理中的膜转运(20个基因)和信号传导(10个基因)。这些注释结果为深入分析低-高质量浓度葡萄糖培养体系中蓝细菌的生长代谢情况提供了有效信息,揭示了在不同质量浓度葡萄糖培养的影响下,蓝细菌可能通过调控特定的基因或代谢通路以适应生存环境中的可变因素,进而改变自身的生物学功能及生长代谢过程。
图 8c、d为KEGG通路富集气泡图,展示了T1、T3组与CK组中蓝细菌的差异表达通路。结果显示,T1组蓝细菌前2个显著差异的代谢通路富集到了光合作用和依赖ATP的ABC转运蛋白,说明碳源葡萄糖的存在影响了蓝细菌的光合作用以及体内的能量运输,光合作用通路总体表现为上调,但依赖ATP的ABC转运蛋白的基因表达水平总体表现下调(表 2),或成为抑制该培养体系对蓝细菌叶绿体内光合色素形成的原因之一。T3组蓝细菌有7个差异显著的代谢通路,显著性从强到弱依次是:氮代谢、光合作用、依赖ATP的ABC转运蛋白、氧化磷酸化、光合生物碳固定、卟啉代谢、乙醛酸和二羧酸代谢。综上所述,这些通路均提示集胞藻PCC 6803在高质量浓度葡萄糖的环境下其光合作用、氧化磷酸化以及碳固定等生命过程的正常进行会被抑制,导致对原本葡萄糖的吸收和利用减少,对生存构成了严重的威胁。
2.1. 不同葡萄糖质量浓度对集胞藻PCC 6803生长的影响
2.1.1. 藻细胞密度
2.1.2. 叶绿素a
2.2. 碳源葡萄糖对集胞藻PCC 6803的代谢影响
2.2.1. 培养液的pH值
2.2.2. 碱性磷酸酶的活性
2.3. 碳源葡萄糖对集胞藻PCC 6803细胞形态的影响
2.4. 转录组学分析
2.4.1. 表达水平差异分析
2.4.2. GO富集分析
2.4.3. KEGG富集分析
-
现如今我国众多河流湖泊面临富营养化的生态问题,了解蓝细菌在水华期间的持续变化情况,对预警与治理蓝细菌水华具有重要的意义[30]。浮游植物优势种/属的演替主要受外界环境条件和物种间相互作用的影响[31]。文献[32]对巢湖浮游植物群落动态变化的研究中提到,夏季的卷曲长孢藻(Dolichospermum circinale)以异形胞固氮方式成为优势蓝细菌,但近年来由于该湖泊氮、磷含量的升高导致非固氮微囊藻的比例开始增长。文献[33]也指出,在洱海稳态转换后,阳光取代水体氮营养成为推动藻类演替的关键因素,完成了由蓝细菌、芽孢杆菌门和隐藻门到蓝细菌、绿藻门和芽孢杆菌门的演替变化。同样地,2012-2022年,吉林省西北部的淡水湖——查干湖浮游植物优势种从硅藻转变为蓝细菌,强调了在浑浊水体中,高NH4+-N和NO3--N含量的比值有利于蓝细菌生长,抑制硅藻生存[34]。由于湖泊干化、盐化,以及水体营养盐含量的增加,岱海优势藻由硅藻和绿藻转变为蓝藻[35-38]。
我国长江三峡库区一级支流澎溪河常年暴发蓝细菌水华,最严重的时期为每年4-5月[39]。截至2022年4月中旬,蓝藻门生物量占浮游植物总生物量的39%,而在水华季后期对表层浮游植物的研究中发现,优势藻种逐渐由光合自养的蓝藻门长孢藻演替为混合营养型甲藻门飞燕角甲藻(Ceratium hirundinella),并且二者的生物量的负相关性有统计学意义;直至2022年5月底,飞燕角甲藻的生物量占浮游植物总生物量的96%以上[40]。除角甲藻体型大且能垂向获取营养的竞争因素外,富营养化水体中溶解性有机质的累积也成为威胁蓝细菌生长以及削弱其竞争营养能力的重要因素。除此之外,三峡库区的大宁河[41-42]、香溪河[41, 43]等一级支流均有甲藻门占优的报道[44]。已知甲藻可以通过鞭毛运动越过温跃层,主动进行垂直运动,以获得下部水体营养,从而获得竞争优势,而蓝细菌因其伪空胞浮力调节结构难以突破温跃层,只能浮在水面,所以在较深的水体中甲藻易繁殖与规模扩张,而在浅水湖泊中通常是蓝藻门之间的藻类相互演替。
在太湖北部的冬春季节,蓝藻门生物量占比接近于0,而隐藻的生物量占比范围为30%~70%;到夏季蓝藻水华频发,其生物量占比范围为60%~87%,而隐藻生物量占比低于10%[45]。依本试验结果可知,随着富营养化程度的加剧,伴随着水体溶解性有机质的不断富集,蓝细菌会逐渐失去生长优势,与此同时,兼养型与异养型微生物的滋生不断消耗水体氧气导致水体缺氧,进而加剧富营养化水体恶化,对此必须加以干涉与治理。本研究通过监测蓝细菌的动态变化,帮助理解水华生消过程中的驱动因子,进而为完善重要水体水华的预警工作提供理论资料。
-
在模拟的封闭试验环境中,铜绿微囊藻(Microcystis aeruginosa)会受到由自身产生的溶解性有机质的逐渐积累所带来的毒害作用而被抑制生长,甚至导致死亡,而异养细菌施氏假单胞菌的存在可以缓解这种毒性,使得两者实现互利共生[46]。卷曲鱼腥藻(Anabaena circinalis)在有机碳源为乙酸钠或葡萄糖时表现出良好的生长特性,但有研究指出鱼腥藻因缺少转体而难以利用葡萄糖[47-48]。本试验选用蓝细菌集胞藻PCC 6803作为研究对象,其产生的蓝细菌水华广泛分布在高校景观水体[49]、富营养水库[50]以及贵州的红枫湖[51]等,作为实验藻种具有易获得的优势。作为一种单细胞蓝细菌,集胞藻的生命周期短,易培养,并且不产生毒素[52]。
蓝细菌细胞对于利用何种有机碳源以及利用程度具有偏爱性。培养试验结果显示,蓝细菌在CK组比在其他组的生长状况好,这说明了蓝细菌在高浓度富有机质环境中不适应。同时,该结论也得到了转录组结果的支持。在葡萄糖质量浓度高达2.1 g/L的营养环境里,PCC 6803的光合作用、氧化磷酸化以及光合生物碳固定通路的差异表达基因表达水平均总体下调,而与氮代谢相关的基因的表达水平均上调,即蓝细菌通过加强氮元素代谢消耗胞内多余能量以减轻光抑制造成的伤害[53],与集胞藻PCC 6803在镉(Cd)胁迫中响应的转录组学结果一致[54];而在较低葡萄糖质量浓度(0.7 g/L)处理下,光合作用通路的基因表达水平上调,依赖ATP的ABC转运蛋白的基因表达水平总体表现下调。因此,低质量浓度有机质可在一定程度上增强蓝细菌的生命活性,但在高质量浓度富有机质水体中,蓝细菌集胞藻的光合作用以及碳固定等细胞和代谢过程被抑制,同时细胞结构被破坏,生物量占比下降,使其不具备较强的竞争力,其优势可能被其他混合营养型的藻类所代替。
蓝细菌虽无叶绿体,但因其类囊体内含叶绿素、叶黄素和藻蓝素等光合色素,从而能够进行光合作用[55, 9]。叶绿素a作为光合作用的重要色素之一,与蓝细菌的光合效率密切相关,也间接影响着蓝细菌的生长。本试验结果显示,在有外源葡萄糖的参与下,营养液中叶绿素a的含量明显减少,这与文献[56]的研究结果相符,即兼养条件下微藻的细胞色素含量普遍降低。同时,研究过程中观察到随着葡萄糖质量浓度的增加,藻细胞密度呈现小幅波动现象,相较于文献[57]的葡萄糖质量浓度(0.9 g/L),本试验葡萄糖质量浓度高出很多,最终试验结果形成鲜明对比。也有学者认为,有些微藻体内可能缺少相应的酶,导致其不具备吸收利用胞外有机质的机能,从而造成藻细胞无法大量繁殖[58],该理论是否可以作为影响本试验结果的因素还有待对代谢学做进一步的研究。同时,由前人的试验结果发现,在葡萄糖耗尽以后,初始质量浓度低的葡萄糖培养液的集胞藻仍能缓慢生长,而初始质量浓度高的葡萄糖培养液中集胞藻的生长速度下降[59]。扫描电镜的图像显示,葡萄糖处理组下的藻细胞均出现凹陷、褶皱等异常生长形态,细胞周围充满胞外多糖,依据文献[60]的研究,推测胞外附着物的存在为营养基质提供了一个从培养基到细胞的有益的向内扩散浓度梯度。
碳源葡萄糖影响着蓝细菌细胞的生化反应以及水体理化性质。在有机质丰富的水体中,藻类光合作用影响CO2缓冲体系[61],从而改变水体pH值,反过来pH值的波动也会影响藻类对有机碳的利用效率。添加葡萄糖后,微藻在兼养条件下呼吸速率均高于光合自养与异养培养[56, 62-63],O2被消耗以及CO2的不断积累是PCC 6803生长液pH值呈现下降趋势的主要原因。水体pH值与水体里有机质的分解、微生物的活动和水生生物的代谢等密切相关[64-66]。同时,微藻的生长由氮、磷营养因子共同限制[67]。在水体环境中,藻类还可以分泌碱性磷酸酶吸收水体中的有机磷[68-69]。碱性磷酸酶活性的表达与蓝细菌生长密切相关,文献[68]的研究表明,碱性磷酸酶编码基因可以被细胞外正磷酸盐抑制,这将导致碱性磷酸酶活性与藻类生长呈负相关。碳源影响着碱性磷酸酶活性,以葡萄糖为碳源时,分解有机质的微生物的酶活性更强[70];同时,在本试验的转录组结果中,两个组别中参与磷酸根离子跨膜转运相关的差异表达基因表达水平均显著下调(p<0.05),表明细胞吸收无机磷的过程受到抑制,从而诱导藻细胞加速分泌碱性磷酸酶,以此克服磷的限制作用。
-
本研究以BG11-葡萄糖作为溶解性有机碳源,对蓝细菌集胞藻PCC 6803的生长状况及生物学过程进行了研究,发现低质量浓度有机质水体可在一定程度上增强蓝细菌的生命活性,但在高质量浓度富有机质水体中,蓝细菌的光合作用以及碳固定等细胞和代谢过程被抑制,并且通过加强氮代谢消耗胞内多余能量以减轻光抑制造成的伤害,叶绿素a的合成受阻,进而影响其正常生长繁殖,甚至对细胞表面结构具有破坏作用。蓝细菌集胞藻所受到的高质量浓度有机质胁迫使其不具备较强的竞争力,其生长优势可能被其他混合营养型的藻类所代替。本研究结果同时也可以为蓝藻水华的治理和水华优势藻演替的研究提供科学资料。