


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
随着人口增长,我国粮食需求持续上升,对耕地要求日益增高,提高农作物产量与实现可持续耕作已成为农业研究的关键课题[1]。近年来,有学者证实了绿肥与秸秆等有机物料还田的增产与生态效益[2]。秸秆是可循环有机物质,还田处理能增强土壤保水保墒,提升土壤有机质含量,减少化肥用量并降低成本。绿肥是传统农业与可持续生态农业的重要部分,因与根瘤菌形成的固氮共生系统,豆科植物成为绿肥选育主力。豆科绿肥生长与翻压处理后会向土壤供氮,助力含氮微生物生长,改善土壤质量,促进作物生长发育,达到增产增收目的。秸秆和绿肥是可利用的生物质资源,将其覆盖或翻压还田,是提高农田生态系统生物质循环效率的有效途径之一,形成“秸秆—作物—土壤”微环境,为作物提供良好的水热条件。同时,秸秆和绿肥中蕴含着大量的氮、磷、钾元素和有机质等养分资源[3],将其还田有助于补充土壤养分,缓解土壤养分失衡等问题。
胞外酶由土壤微生物于植物根部合成,并催化土壤中重要的生化反应。土壤微生物将胞外酶释放出来,这些酶按照严格的比例通过生物信号途径调节土壤养分循环以及微生物的群落组成,因此胞外酶的化学计量比亦可作为微生物获取和分配碳、氮、磷以及其他养分的预测指标[4-5]。近年来,土壤碳、氮、磷获取酶化学计量比与微生物营养策略及养分限制间的关联,已成为土壤微生物生态学研究的热点[6-7]。Sinsabaugh等[8]基于全球范围大尺度Meta分析提出ln(BG)∶ln(NAG+LAP)∶ln(AP)的化学计量比值接近1∶1∶1,被限制在一个相当窄的范围内。诸多研究已经证实田间有机物投入能够改变土壤胞外酶化学计量比。土壤碳、氮、磷获取酶亦相互影响,调节土壤微生物养分限制[9]。在一项草原生态系统氮添加试验中发现,ln(BG)∶ln(NAG+LAP)和ln(BG)∶ln(AP)比值随着氮素的增加而增加,表明在环境中氮素养分过剩的情况下,相对于氮获取酶和磷获取酶,土壤微生物对碳获取酶的投资增多,以获得更多的碳,缓解生长过程中碳养分的限制[10]。
我国西南丘陵地区受自然条件限制,“旱三熟”种植模式成为本区旱作农田普遍采用的分厢套作多熟复种的主要增产模式。这种复种方式用地强度大,进而易引发土地用养失衡、土地质量下降等诸多问题,因此亟待探索可持续耕作措施以缓解土地用养失衡与土地质量下降等现象。目前,国内外学者在秸秆覆盖、绿肥种植等保护性耕作对土壤肥力、微生物多样性影响等方面均展开了较多研究,而现有研究主要局限在稻田以及一年一熟或者两熟种植区,对西南丘陵地区秸秆绿肥覆盖还田的相关研究较少[11]。因此,本研究以西南地区“蚕豆/玉米/甘薯” “旱三熟”套作模式中的蚕豆为研究对象,将秸秆覆盖和紫云英绿肥覆盖相结合,研究不同覆盖措施对土壤养分、胞外酶活性及其化学计量比和作物产量的影响,旨在为优化农田管理提供科学依据。
全文HTML
-
试验地点位于重庆市北碚区西南大学紫色土肥力与肥料效益监测站(29°51′ N,106°27′ E),海拔244 m。该地区属于亚热带季风气候,年均气温约18.2 ℃,年均降雨量约1 100 mm,降雨主要集中在5-9月。年均太阳总辐射量87 108 kJ/cm2,年均总日照时数1 276.7 h。试验地土壤为旱地紫色土,地势平坦,土壤肥力均匀。
-
试验种植模式为西南旱地农田“蚕豆/玉米/甘薯”三熟制套作模式,蚕豆(Vicia faba L.)供试品种为重庆本地品种‘小青蚕豆’,玉米(Zea mays L.)供试品种为‘先玉1171’,甘薯(Ipomoea batatas L. Lam.)供试品种为‘渝红心薯98’。基于2018年开始的长期定位试验,本研究试验期为2022年11月22日至2023年5月14日,共设计4个处理:无覆盖(CK)、秸秆覆盖(S)、紫云英覆盖(M)以及秸秆与紫云英协同覆盖(SM)。基于团队前期研究成果,本试验设置蚕豆、玉米秸秆覆盖量为4 500 kg/hm2[3-4],紫云英覆盖量为2 500 kg/hm2。具体操作方法如表 1所示。
本试验采用单因素随机区组试验设计,每个处理设置3个重复,共12个小区,每个小区面积为8 m×4 m=32 m2,每个小区按长4 m、宽1 m分成8个条带。蚕豆和紫云英同时于2023年11月22日以间套作方式播种,蚕豆每个条带种植2行,每行12窝,每窝2株,于2023年4月5日收获紫云英并同时种植玉米,于2023年5月14日收获蚕豆并同时种植甘薯。蚕豆生长期秸秆覆盖和紫云英覆盖时间分别为2022年11月23日和2023年4月8日。施用复合肥(含N 15%、P2O5 15%、K2O 15%)750 kg/hm2和尿素150 kg/hm2作为基肥,在玉米与甘薯移栽时一次性施入,蚕豆播种时不施肥,其他田间管理均按常规措施进行。
-
分别于蚕豆苗期(2022年12月31日)、分枝期(2023年1月31日)、花期(2023年3月4日)、结荚期(2023年4月1日)、成熟期(2023年4月29日)进行土壤样品采集。每次进行土壤取样时选取不同处理小区内生长状况一致的蚕豆3穴,用工具挖出蚕豆植株的完整根系,抖落多余土壤,分离出植株根部0~20 cm土层的根际土壤(附着在蚕豆根系0~10 mm处),收集土壤约600 g,将同一小区的3穴土壤样品混合均匀后作为一个样品装入密封袋中,立即置于冰盒内冷藏带回实验室。将取回的鲜土挑出其中石块和杂草根系,过2 mm筛混合均匀后分成两部分:取用100 g储存于4℃冰箱,用于土壤微生物生物量养分、土壤胞外酶活性试验测定;剩余部分自然风干后,过2、1、0.25、0.149 mm筛后用于测定土壤碳、氮、磷、钾含量。
-
取用苗期、分枝期、花期、结荚期以及成熟期土壤样品进行土壤养分测定。使用pH计测定土壤pH值,水土比为2.5∶1;烘干法测定土壤含水率(SWC);TOC-LCPH自动分析仪(日本)测定土壤总有机碳(TC)含量;浓H2SO4消解-凯氏定氮法测定土壤全氮含量(TN);NaOH熔融-钼锑抗比色法测定土壤全磷含量(TP);NaOH熔融-火焰光度计法测定土壤全钾含量(TK);碱解扩散法测定土壤碱解氮(AN)含量;钼酸盐-抗坏血酸比色法测定土壤有效磷含量(AP);乙酸铵浸提-火焰光度计法测定土壤有效钾含量(AK)[12];氯仿熏蒸浸提法测定土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)含量[13]。
-
取用蚕豆成熟期土壤样品进行胞外酶活性测定。采用荧光微孔板检测技术测定β-1,4-葡萄糖苷酶(BG)、纤维二糖水解酶(CBH)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)、亮氨酸肽酶(LAP)和酸性磷酸酶(AP)5种胞外酶活性,不同酶种类、测定活性所用标准品和底物如表 2所示,相关酶活性计算参照German等的研究[14]。
-
蚕豆收获期,在每个小区内随机选择20株蚕豆记录每株豆荚数和每荚籽粒数,随后将收获后的蚕豆脱粒后风干称质量,计算百粒质量和籽粒产量。
式中:a为籽粒产量(kg/hm2);b为每公顷株数(株);c为每株荚数(个);d为每荚粒数(粒);e为百粒质量(g)。
-
使用IBM SPSS Statistics 25进行单因素方差分析(ANOVA),并采用Duncan post hoc检验不同生物覆盖处理之间土壤理化性质、胞外酶活性的差异。通过Origin 2023 pro进行图形绘制。
1.1. 试验地概况
1.2. 试验设计
1.3. 样品采集
1.4. 土壤指标测定
1.4.1. 土壤养分衡量指标的测定
1.4.2. 土壤胞外酶活性的测定
1.5. 产量测定
1.6. 统计分析
-
生物覆盖过程中,整个生育期土壤pH值为5.80~6.16(表 3),生物覆盖与无覆盖差异无统计学意义,4个处理中,紫云英覆盖处理下土壤pH值相对更小;各个生育期各处理之间含水率变化趋势保持一致,均呈“W”形变化(表 4),各处理之间差异无统计学意义,整个生育期内含水率变化范围为12.21%~25.68%,在花期SM处理下的含水率达到最高。
如表 5所示,蚕豆成熟期S处理下的土壤全钾和碱解氮显著高于CK、M和SM处理;S处理下总有机碳、全氮、速效钾含量高于其他处理但差异无统计学意义;土壤全磷的S和M处理高于CK处理;土壤有效磷含量表现出SM处理高于其他处理但差异无统计学意义。总体而言,S处理在提高土壤全钾和碱解氮含量方面有显著效果,虽然总有机碳、全氮、全磷、有效磷和速效钾含量没有显著提升,但整体含量处于较高水平,SM处理的有效磷和速效钾含量处于较高水平,对提高土壤养分有一定作用,不过其全钾和碱解氮含量不如S处理。SM处理的有效磷和速效钾含量处于较高水平,对提高土壤养分有一定作用,不过其全钾和碱解氮含量不如S处理。
-
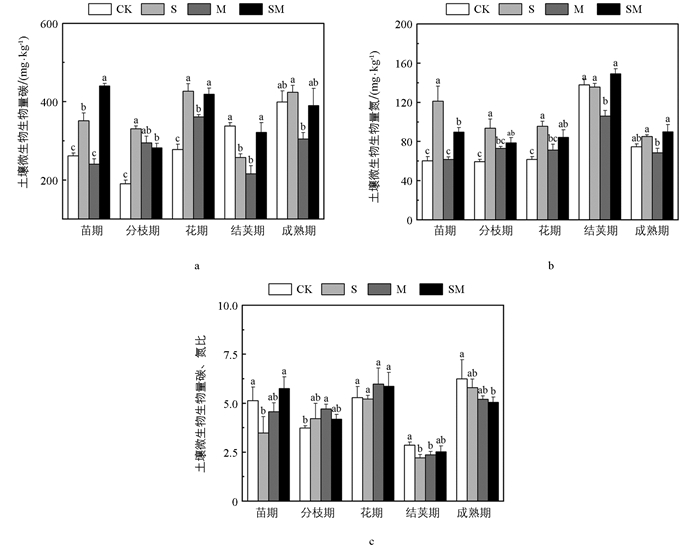
图 1a是土壤微生物生物量碳含量的动态变化情况,不同处理和不同时期的土壤MBC存在显著差异。苗期,SM处理下的土壤MBC含量显著高于其他处理,S处理显著高于CK、M处理;分枝期,S、M、SM处理显著高于CK处理;花期,覆盖处理的土壤MBC含量均显著高于无覆盖处理;结荚期,CK处理与SM处理显著高于S和M处理;成熟期,S处理显著高于其他处理。土壤MBN含量方面,在苗期、分枝期和花期,S和SM处理显著高于CK处理;在结荚期与成熟期,M处理显著低于S和SM处理(图 1b)。
土壤微生物生物量碳、氮化学计量比(MBC∶MBN)范围为2.21~6.29(图 1c)。苗期,S处理下的MBC∶MBN最小,为3.49;分枝期,除S处理的MBC∶MBN值上升外,其余处理均略微下降,其中CK处理下降幅度最大;结荚期,各处理MBC∶MBN值均较花期大幅下降,S处理和M处理显著低于CK处理;成熟期时,S、M、SM处理下的MBC∶MBN值均低于CK处理。
-
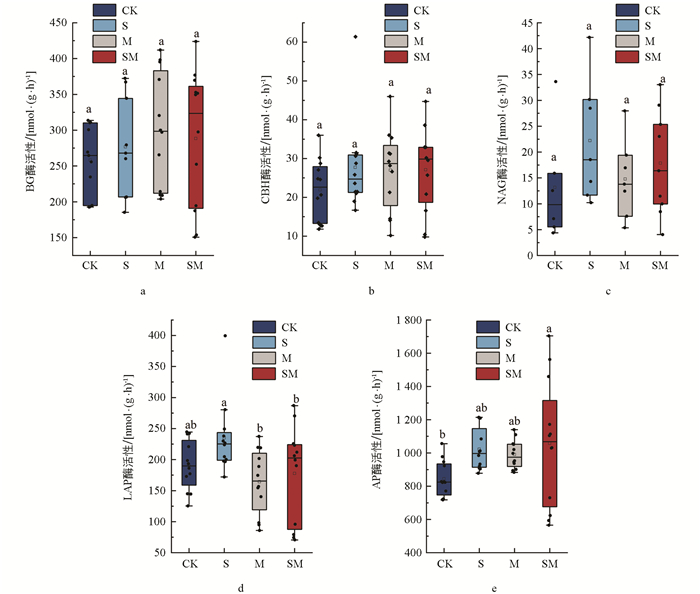
生物覆盖改变了蚕豆成熟期土壤C、N和P获取酶活性(图 2)。各处理之间BG酶、CBH酶以及NAG酶活性差异并不明显。不同处理的BG酶活性与CBH酶活性大小为:M>SM>S>CK;NAG酶活性无显著差异,其大小顺序为:S>SM>M>CK。LAP酶活性方面,S处理显著大于其他处理;AP酶活性的SM处理显著高于CK处理,AP酶活性大小为:SM>S>M>CK,其中SM处理显著高于M和SM处理。综合来看,S、SM处理在增加土壤C、N和P获取酶活性效果方面表现最佳。
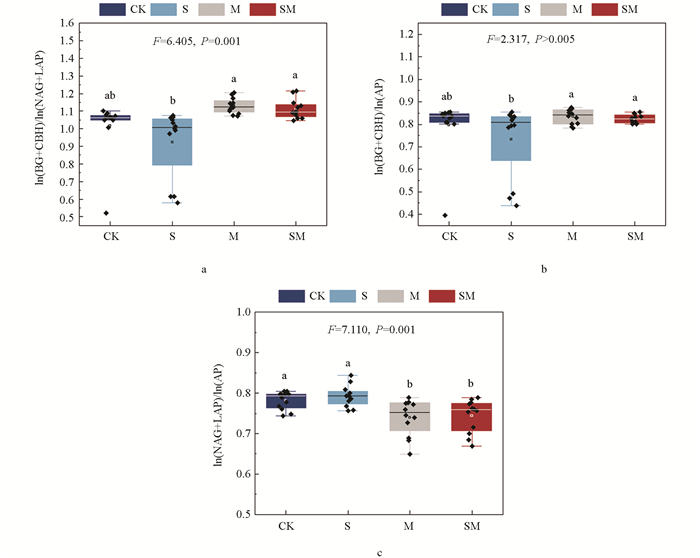
生物覆盖显著改变了土壤胞外酶化学计量比(图 3),土壤ln(BG+CBH)∶ln(NAG+LAP)值和ln(BG+CBH) ∶ln(AP)值均表现为M和SM处理显著高于S处理;CK处理与其余处理相比,均无显著差异。M和SM处理ln(NAG+LAP)∶ln(AP)值显著低于CK处理和S处理。
-
如表 6所示,CK、S、M、SM处理下的蚕豆平均产量分别为1 267.41、1 522.60、1 489.72、1 601.56 kg/hm2,S、M和SM处理产量相较于CK处理显著提高了20.13%、17.54%、26.36%。覆盖处理对蚕豆产量的增加强度排序为:SM>S>M>CK。综上所述,生物覆盖处理能显著增加蚕豆的产量,其中秸秆和紫云英协同覆盖效果最好。
2.1. 不同覆盖处理下土壤理化性质的变化
2.2. 不同生物覆盖处理下土壤微生物生物量碳、氮含量及碳、氮比的变化
2.3. 不同生物覆盖下成熟期土壤胞外酶活性变化及其化学计量学特征
2.4. 蚕豆产量
-
土壤水分含量与降水量、水分蒸发量以及植物水分利用效率密切相关。有机物料覆盖还田后在地表土壤与大气之间形成隔层,延缓了土壤毛管水向表层土壤的运移,从而减少水分蒸发量。此外,秸秆等有机物料能够吸收一定的水分,延长了自然降水在土壤表层的滞留时间[15-16]。本试验中,生物覆盖处理下的土壤含水率整体上高于无覆盖处理,表明生物覆盖提高了土壤水分含量,这与前人研究结果一致[16-17]。
作物秸秆和绿肥作为一种优质的有机肥源,富含N、P、K等营养元素和有机质等养分资源,还田后能够有效地为土壤和后茬作物提供养分资源[17]。本试验结果显示,秸秆和紫云英还田均能提高土壤的有机碳、全氮含量,但其含量变化趋势不尽相同。具有秸秆覆盖的处理土壤有机碳含量在蚕豆成熟期高于无秸秆覆盖,S处理有机碳含量最高,SM处理则仅次于S处理。土壤速效养分反映了土壤中相应元素的供应能力,速效养分能够被植物直接吸收利用,其含量对作物的生长发育起着重要的作用。在本试验中生物覆盖处理显著增加了土壤硝态氮含量,有效磷和速效钾的含量略微提升,这与前人的研究结果一致[18-19]。在植物体内的磷元素约有40%~60%以水溶态或弱酸溶解态的形式存在,在秸秆还田早期能够迅速进入土壤中,其余部分以磷酸、核酸以及磷酸肌醇的形式参与细胞壁、核酸等的构成,在一定酸性条件下,需要依靠微生物进行降解[20-21]。还田有机物料中的磷释放率与自身磷含量呈正相关。有研究表示,水分充足条件下,植物残体中的有机磷能被迅速溶解释放[22]。对于土壤全磷含量来说,在蚕豆成熟期没有表现出差异,其中SM处理下全磷含量最低。本试验中,由于2023年的降水量较低,土壤中水分含量较低,秸秆中的有机磷无法释放进入土壤,从而引起土壤全磷含量的下降。此外,本研究发现,生物覆盖能够有效地提高土壤的肥力。因此,为了进一步得到更加明确的结论,并开展长期的定位试验,研究秸秆与紫云英覆盖条件下土壤全量养分及其化学计量比是十分必要的。
土壤微生物生物量与土壤肥力密切相关,是评价土壤质量的重要生物学指标。有机物料还田后能为微生物生长繁殖提供丰富的营养物质,从而增加微生物生物量碳、氮、磷[23]。本研究发现,在蚕豆成熟期S、SM处理下的微生物生物量碳含量高于CK处理,这与邢文超等[24]的研究结果一致。对于微生物生物量氮含量,在苗期、分枝期、花期和成熟期,S、SM处理均高于CK处理,说明生物覆盖能够提高土壤微生物氮的含量。根据“微生物化学计量平衡理论”,不同种类有机物料共同还田可以形成适宜的C∶N∶P,成为更利于微生物利用的养分组合,促进微生物生长和秸秆腐解[25]。在农业生态系统中,耕作措施、季节、施肥、投入外源有机物质都会影响微生物生物量化学计量特征,从而使得土壤化学计量学特征波动范围较大。Cleveland等[26]基于全球范围得出MBC∶MBN∶MBP保持在约60∶7∶1的比例;刘晗等[27]通过施加不同绿肥,发现红壤幼龄橘园MBC∶MBN范围为10.00~22.68;余顺平等[28]研究得到在不同水平的施肥处理下MBC∶MBN波动范围为4.75~7.62。本研究也验证了这一点,蚕豆成熟期MBC∶MBN为4.52,偏离全球平均水平,说明本研究区域受C限制高于N限制。
土壤胞外酶活动是将还田有机物料腐解反应与微生物代谢利用营养物质相联系的关键生物学过程。其活性变化与土壤养分含量密切相关,在调控土壤养分地球生物化学循环中发挥着重要作用。此外,酶对环境生态因子具有高度的灵敏性,因此酶常常被作为反映植物—土壤—生物系统波动的重要指标。孙娇等[29]将有机物质还田发现可以增加土壤BG、CBH、NAG、LAP酶活性,这与本研究结果相似。一般认为土壤酶活性与土壤养分含量呈正相关,生物覆盖中的秸秆与紫云英还田,一方面可以作为外源碳为土壤微生物活动提供充足的碳源;另一方面,外源碳的投入使土壤微生物引发激发效应,刺激土壤微生物分泌胞外酶分解有机质,加速土壤养分循环。土壤胞外酶C∶N∶P代表了微生物C、N、P营养需求与养分资源供应之间的平衡,全球尺度范围内土壤胞外酶C∶N∶P约为1∶1∶1[8]。Waring等[4]基于较低的土壤酶C∶P和N∶P比例,以ln(BG)∶ln(AP)和ln(NAG)∶ln(AP)为代表,认为土壤P是限制热带生态系统养分循环的元素。本研究中,2023年土壤胞外酶C∶N和N∶P为分别为0.55、0.76,因此土壤N和P元素是限制本研究区域的主要元素。本试验发现,S、SM处理相较于CK处理可以缓解N、P限制。
前人报道称绿肥种植、秸秆还田能够提高作物的产量[30]。与无覆盖对照相比,单独覆盖秸秆、紫云英以及两者共同还田的蚕豆籽粒产量显著提高了17.54%~26.36%,以秸秆与紫云英共同还田增产效果最佳,秸秆单独还田次之。3种生物覆盖方式中,秸秆与紫云英共同还田下蚕豆产量最高。可能原因为:一方面,与秸秆、紫云英单独还田相比,紫云英与秸秆共同还田能延长养分的有效性,协调秸秆、紫云英还田后养分释放与作物养分吸收之间的关系[31];另一方面,高C∶N的秸秆与低C∶N的紫云英共同还田下养分组成更加合理。
-
生物覆盖处理能够提高土壤肥力,其中对于土壤速效养分含量来说,S、SM处理相较于CK处理的效果显著;生物覆盖显著提高了土壤微生物生物量碳、氮含量,从各个生育期来看,S、SM处理的土壤微生物生物量碳、氮含量显著高于CK处理;生物覆盖处理下的蚕豆产量显著提高,以SM处理下的蚕豆产量最高。综上所述,农田生物覆盖措施能够改良土壤理化性质,提升土壤获取酶活性。相较于其他处理,S、SM处理在提高土壤肥力、胞外酶活性以及蚕豆产量方面更加具有优势,可作为优化西南地区旱作农田管理的技术措施。