


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
裂果是柑橘生产中一种常见的生理性病害,是由于果皮与果肉生长不一致造成的. 柑橘果肉细胞迅速膨大,果实的膨胀压增大,胀破果皮,从而造成果实开裂. 宽皮柑橘、甜橙、柚等品种常有裂果发生,裂果率达20%~30%,严重时可高达70%[1];另外,红江橙[2]、锦橙[3]、脐橙[4]、温州蜜柑[5]等品种果实亦有报道裂果的发生. 据统计,温州蜜柑裂果率可达20%~40%,锦橙裂果率可达30%~40%,脐橙不同品系裂果率一般在2%~14%[6]. 裂果对果实的商品价值和销售特性造成了严重的负面影响,给柑橘产业带来极大的经济损失.
柑橘裂果在果实膨大期至果实成熟期均有发生[7],但不同品种的裂果类型和裂果发生时间有较大差异. 柑橘裂果主要分为外裂和内裂两种类型,二者都是一个渐变的过程. 早期内裂发生在果实的内部,前期内裂果与正常果没有明显差异,不易察觉;中后期果实内裂进一步向外扩展,在果皮表面出现明显的凹凸不平,严重时造成果皮的开裂,如纽荷尔脐橙[8]和早香柚的内裂[9];外裂是从果顶或果面开裂,由于角质层受到挤压而出现裂缝,由外向内发展,细胞逐渐变形、破裂,最终海绵组织破裂,直至外果皮断裂,形成明显裂口[10],如朋娜脐橙的裂果[11].
柑橘果实的生长发育受果实含水量的影响,而果实含水量又受空气湿度和土壤水分的双重调节[12],其中土壤水分直接影响果实含水量,是造成裂果的关键因素. 已有研究表明,土壤水分的剧烈变化和水分不合需求均能引发裂果[13]. 剧烈的土壤水分变化会加剧裂果的发生,如改良橙在果实生长过程中,久旱后遇骤雨,裂果现象的发生更为严重[14]. 水分胁迫也会造成柑橘果实裂果,Lima[15]研究发现,柑橘树体在土壤水分胁迫下会使根系受损,降低了吸水能力,进而导致果皮生长受到抑制,但果肉生长能力不变,果皮生长速率跟不上果肉组织生长速率,最终果肉组织膨压导致裂果. 赖呈纯等[16]研究表明,采用适时喷灌的措施,使土壤含水量保持较均衡并在临界值之上,可有效防止茂谷柑裂果;朱潇婷等[5]研究发现在干旱处理下,较少的灌水变化幅度会加重温州蜜柑的裂果. 水分是果树生长过程中矿质元素吸收和运输的载体,而矿质元素的丰缺影响着果实的发育和代谢进程. 周利[17]研究发现,在水分胁迫条件下纽荷尔脐橙和温州蜜柑果实的钾、钙、镁元素的吸收随土壤水分减少而降低;许建楷等[18]报道红江甜橙的正常果实钙含量高于裂果果实,并且发现果实的裂果率与钙含量呈负相关;春季或果实早期施用钾肥可以促进果皮发育,增加果皮厚度,增强果实的抗裂能力,减少收获前果实的开裂[19].
甘平(Citrus Kanpei)是由西之香和椪柑杂交而来的柑橘新品种,具有树势强、果型大等特点,风味独特、糖酸适中、耐贮藏,是一个较有潜力的中晚熟杂柑品种. 但该品种裂果率较高,如遇气候条件不利的年份或栽培措施不到位,裂果率可高达七成,该难题制约了甘平品种的大面积推广[20]. 重庆地区夏季高温少雨,降雨时间分布不均,甘平裂果尤为严重[21]. 柑橘裂果与水分密切相关,但关于甘平裂果防治研究中鲜有控制含水量及变化对减轻裂果的报道,水分对果实开裂内部机理的调控机制研究仍需深入. 本试验利用水肥一体化智能灌溉系统,精确控制土壤含水量及水分变化幅度,探究果实内部生理变化及其与裂果的相关性以及有效防控甘平裂果发生的最佳土壤含水量,旨在为甘平优质高效栽培提供技术支撑.
全文HTML
-
试验于2021-2022年在西南大学柑桔研究所试验地进行,供试验的柑橘为6年生香橙砧甘平(Citrus Kanpei),果园施肥和病虫害控制统一按常规进行. 试验果园土壤基本肥力状况为:土壤pH值为5.41,有机质17.75 g/kg,碱解氮106.19 mg/kg,有效磷61.84 mg/kg,速效钾173.82 mg/kg,有效钙1 853 mg/kg,有效镁37.46 mg/kg.
-
前期田间调查结果显示甘平裂果与土壤水分变化密切相关,因此于2021年对土壤水分变化幅度进行精准调控,共设定5个土壤含水量变化阈值,分别是5%(△SWC5,土壤相对含水量90%~95%),20%(△SWC20,土壤相对含水量75%~95%),35%(△SWC35,土壤相对含水量60%~95%),50%(△SWC50,土壤相对含水量45%~95%)和自然降雨(对照,CK),利用水肥一体化灌溉设备监测土壤含水量,低于设定阈值后自动开启灌溉系统,土壤含水量达95%时停止灌溉.
-
于2021-2022年探究减少甘平裂果的最佳土壤相对含水量,试验共设5个处理,分别将土壤相对含水量设定为45%(±5%,SWC45),60%(±5%,SWC60),75%(±5%,SWC75),90%(±5%,SWC75)和自然降雨(对照,CK). 自第二次生理落果后开始处理,控制土壤相对含水量保持在5%上下限范围内. 土壤相对含水量的测定使用土壤温湿度传感器实时监控.
树体根系分布深度在30~50 cm,每棵树在树冠滴水线放置4个滴头,每组处理安装3个土壤温湿度传感器相互校正控制土壤含水量,传感器探头埋于滴水线35~40 cm深处. 试验期间种植区外围开沟,沟深40 cm,垄上铺设防渗膜导出雨水. 两组试验中每处理分别选取9株树,每3株树为一个重复,共计3个生物学重复;第一组试验仅于2021年进行裂果统计,第二组试验于2021-2022年进行裂果统计且分别采集果实及叶片进行生理指标测定.
-
分别于盛花后75 d(6月5日)、盛花后90 d(6月20日)、盛花后105 d(7月5日)、盛花后120 d(7月20日)、盛花后135 d(8月4日)、盛花后150 d(8月19日)、盛花后165 d(9月3日)、盛花后180 d(9月17日)在树冠四周随机采集20个大小一致的果实,置于带冰袋的泡沫盒中,带回实验室. 用去离子水洗净擦干,使用游标卡尺测量果实赤道部横径和纵径,然后在赤道部横切,用游标卡尺测定果皮厚度,果实硬度使用GY-4数显硬度计测定,以硬度计穿刺果实赤道面果皮的力表示,单位为N.
在果实裂果高峰期(8月4日),分别采集各处理下的已裂果和未裂果各30个,分为两组,一组测定果实性状后,将果皮果肉分离,于105 ℃杀青30 min,然后80 ℃烘干至恒质量后研磨成粉用于矿质元素的测定;另一组将果皮和果肉分离,取果皮,混样后用锡箔纸分装,液氮速冻,-80 ℃冰箱保存,用于测定细胞壁物质、水解酶活性、活性氧含量、抗氧化酶活性. 同时采集各处理下当年成熟叶片若干,分为两组,一组立即测定水分指标;另一组于105 ℃杀青30 min,然后80 ℃烘干至恒质量后研磨成粉用于矿质元素的测定.
-
1) 裂果统计. 在生理落果期结束后,记录每株树的果实总数量,自开始出现裂果时观察并记录裂果果实个数,以后每隔15 d记录一次,每次数据采集结束后摘除本次裂果(同时摘除病果),直至果园不再出现裂果,裂果率=(裂果个数/总果数)×100%.
2) 水势测定. 采用露点水势测定系统(美国Wescor Inc公司)测定叶片及果皮水势.
3) 叶片与果皮相对含水量的测定. 相对含水量=(FW-DW)/(TW-DW)×%,式中FW为组织鲜质量,DW为组织干质量,TW为组织浸水4 h后测定的饱和质量[22].
4) 叶片光合指标的测定. 于裂果高峰期时选择晴朗天气,采用3051D便携式光合仪(浙江托普云农公司)和Handy PEA(英国Hansatech公司)在每次采样前原位测定树体东南西北方向各2片当年生春梢叶片的净光合速率(Pn)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr).
5) 细胞壁物质含量的测定. 细胞壁物质的提取和分离参照Brummell等[23]、Toshihiro A等[24]及魏建梅[25]的方法,分别提取得到总果胶、可溶性果胶、半纤维素以及纤维素;不同形态的果胶质量分数采用咔唑—硫酸比色法测定,半纤维素质量分数通过蒽酮比色法测定.
6) 细胞壁水解酶活性的测定. 纤维素酶(CL)活性的测定参照Georg等[26]的方法;果胶酶(PG)活性的测定参照Biz等 [27]的方法.
7) 丙二醛(Malondialdehyde,MDA)、超氧阴离子(Superoxide anion,O2-)和过氧化氢(Hydrogen peroxide,H2O2)摩尔质量的测定. MDA摩尔质量测定参照吴会会等[28]的方法,O2-和H2O2的摩尔质量测定参照王爱国等[29]的方法.
8) 超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT)、过氧化物酶(Peroxidase,POD)活性的测定. 细胞壁代谢酶的测定参照刘欢[30]的方法.
9) 矿质营养元素的测定. 将烘干后的组织粉碎过筛,用H2SO4-H2O2法消煮制备成待测液,分别用半微量凯氏定氮法、钼锑抗比色法、火焰光度法测定N,P,K含量;用HNO3-HClO4消煮制备待测液,使用电感耦合等离子体光谱仪测定Ca,Mg含量,元素测定参照鲍士旦[31]的方法.
10) 气象数据采集. 2021年与2022年的气象数据来源于美国国家海洋和大气管理局(NOAA),包括6-9月各月的主要气象参数[32](降雨量、平均气温、最高气温、最低气温、日照时数、相对湿度)共6项.
-
采用Microsoft Excel 2019进行数据统计分析;采用SPSS 25.0进行Duncan's差异显著性检验;采用Origin 2021进行绘图,最终结果以两年统计数据的均值显示.
1.1. 试验材料
1.2. 试验设计
1.2.1. 土壤水分变化幅度的设置
1.2.2. 土壤相对含水量精准控制设置
1.3. 采样与指标测定
1.3.1. 样品采集
1.3.2. 指标测定
1.4. 数据分析
-
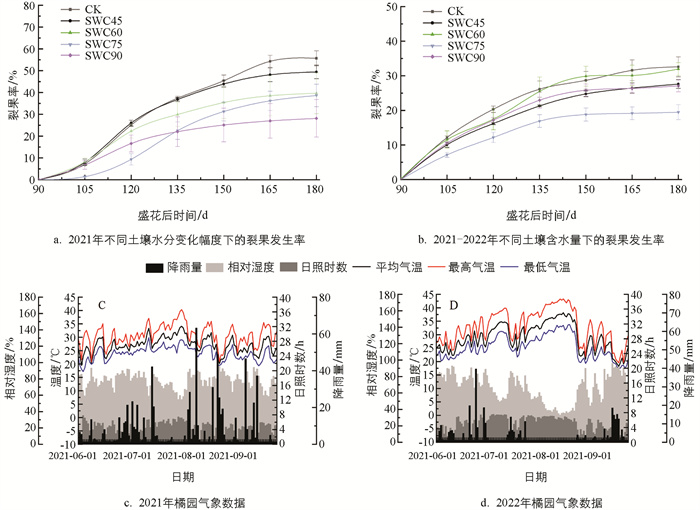
以自然降雨作为对照,采用水肥一体化灌溉设备控制土壤湿度变化幅度,统计各处理下裂果的发生情况(△SWC5,△SWC20,△SWC 35和△SWC 50). 图 1a结果显示,随着土壤水分变化幅度增大,甘平裂果率显著升高. 其中,△SWC35和△SWC50两个处理与CK甘平裂果发生率相近,30 d内均达到20%以上,△SWC5处理的甘平裂果率最低,30 d内低于10%. 当土壤水分变化幅度高于20%时,果实裂果显著增多,土壤水分变化幅度在5%时,极显著降低了果实裂果率.
由图 1b可知,甘平裂果高峰期出现在盛花后120~150 d,即果实膨大期(7月下旬至8月上旬),与对照相比,SWC75处理显著降低了甘平果实裂果发生率,说明土壤含水量在75%能有效预防裂果.
对柑橘园所在地区2021和2022年两年柑橘快速生长期气象数据的分析发现,在柑橘果实快速膨大时期总体呈现高温少雨,干旱突出的特征. 6-8月降雨量少且分布不均匀,连续降雨日短,空气湿度变化大,多为少量降雨后干旱再降雨,高温干燥天气长. 重庆地区7-8月处于伏旱季节,持续高温寡雨,相对湿度低,日照时数长,在自然降雨后,果实裂果量剧增(图 1c、图 1d).
-
裂果与正常果实生理指标测定显示,二者的果形指数差异不明显;正常果的果皮相对含水量低于裂果但差异不明显;正常果果皮水势显著低于裂果. 此外,正常果的果皮厚度和果皮硬度分别比裂果高16.96%和18.19%(表 1).
果皮细胞壁成分分析发现,裂果果皮中总果胶和纤维素的质量分数显著低于正常果果皮,分别低6.50%和14.01%,而可溶性果胶与半纤维素的质量分数低于正常果皮但差异不显著;裂果果皮中果胶酶和纤维素酶的活性显著高于正常果(表 2).
果皮活性氧代谢分析显示,裂果果皮中O2-和H2O2的摩尔质量都显著高于正常果皮,分别高26.69%和44.90%;开裂果皮中MDA摩尔质量略高但差异不显著;裂果果皮中SOD,CAT和POD的活性显著低于正常果,分别低16.49%,19.10%和19.96%(表 3).
-
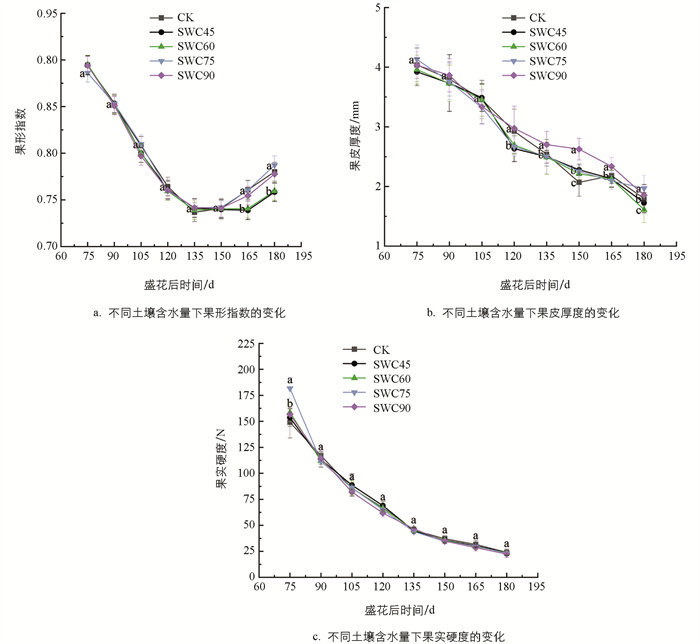
在各处理条件下,甘平果实果形指数自盛花期75 d后均不断减小(图 2a). 在盛花后135~150 d时达到最低,约为0.74,此后果实横径发育减缓,果形指数逐渐上升,至膨大期结束,各处理果形指数基本维持在0.76左右.
在整个果实发育期,75%和90%土壤相对含水量处理下果皮厚度高于其他各处理(图 2b). 盛花后75 d,甘平果实快速膨大,果皮厚度持续变薄,各处理下的果皮厚度变薄的速率总体上呈现出“快—慢—快”的变化趋势;盛花后135 d,果皮厚度降低的趋势逐渐平缓,盛花后180 d;果实进入转色期,SWC60和SWC45处理的甘平果皮厚度较低,SWC75处理的果皮最厚,但各组差异不显著.
随着果实的发育,果实硬度一直降低(图 2c),盛花后75~135 d,硬度降低速率最快;盛花后135 d,果实硬度缓慢降低,膨大期结束果实硬度基本维持在30 N左右,各组处理甘平果实硬度变化基本一致,但不存在显著差异.
-
由表 4可知,土壤含水量在SWC45和SWC60时,果皮的水势降低,SWC75和SWC90处理的果皮水势显著高于CK和其他处理,其中,SWC90处理下的水势略低于SWC75. 结果显示,果皮相对含水量随着土壤相对含水量的升高而升高,CK果皮的相对含水量为60.26%,SWC45和SWC60处理下的果皮相对含水量与CK差异不显著,SWC75和SWC90处理下的果皮相对含水量显著高于CK.
-
从统计结果可以看出,SWC75和SWC90处理下果皮总果胶的质量分数显著高于CK和其余处理,分别为263.11 mg/g和213.90 mg/g;SWC75处理下果皮可溶性果胶质量分数最低且显著低于其余处理;果皮纤维素质量分数在SWC75处理下最高,为272.81 mg/g,SWC45,SWC60和SWC90处理果皮纤维素质量分数与CK无明显差异;果皮半纤维素的质量分数在各处理间不存在显著差异,但SWC75处理的半纤维素质量分数最高,为148.46 mg/g. 分析果皮中两种主要细胞壁水解酶在不同土壤水分条件下的活性差异,结果显示,SWC45处理下果胶酶活性最高,且显著高于其余处理,SWC60和SWC90处理的果胶酶活性低于CK但无显著差异,SWC75处理果胶酶活性最低且显著低于其余各处理;纤维素酶的活性在SWC45处理下最高,其余4组处理间纤维素酶活性无明显差异,SWC75处理下纤维素酶活性最低,为0.56 mg/h/g,与CK相比差异不显著,但显著低于SWC45(表 5).
-
研究结果表明,在SWC60处理下果皮中O2-的摩尔质量最高且显著高于其余各处理,SWC75和SWC90处理下果皮O2-摩尔质量显著低于CK,分别为92.16 nmol/g、106.30 nmol/g;SWC60,SWC75 SWC90处理下的果皮过氧化氢摩尔质量显著低于CK;SWC75和SWC90处理果皮的MDA摩尔质量显著低于CK,其中SWC75处理的MDA摩尔质量最低,为5.22 nmol/g. 分析不同土壤水分条件下甘平果皮中抗氧化酶的活性,结果表明,各处理下SOD的活性都显著高于CK,其中SWC75处理下的果皮SOD酶活性最高,为972.01 U/g;在SWC75处理中,果皮CAT活性为459.56 μmoL/min/g,显著高于对照;SWC45,SWC75和SWC90处理的POD活性显著高于对照,其中SWC90处理最高,SWC75处理次之(表 6).
-
SWC45,SWC60和SWC75中N含量与土壤相对含水量呈正比,SWC75处理下的果皮和果肉中N含量最高,SWC90处理的果皮与果肉中N含量同样有所降低. SWC45和SWC60处理果皮中的P含量显著低于其余处理,SWC45处理果肉中的P含量最低. SWC75处理下的果皮与果肉中K含量也显著高于其余处理. 在果皮和果肉中SWC75的Ca含量最高,且果实膨大期果皮钙含量高于果肉. 此外,随土壤含水量的增加,果实对Ca的吸收增加,但当土壤相对含水量达90%时,叶片和果实对Ca的吸收降低. 果皮中不同水分处理下Mg含量不存在显著差异,果肉中的Mg含量在SWC45处理下含量极低,仅为0.69 g/kg,显著低于其余处理,与CK相比低45.24%,SWC75处理下果肉Mg含量最高,且显著高于其余处理(表 7).
-
由表 8可知,在不同土壤水分下,叶片与果皮的水势差异趋势一致,土壤含水量降低时,叶片水势均降低,SWC75和SWC90处理的叶片水势显著高于CK和其余处理,其中,SWC75处理下的水势最高,SWC60处理下的水势与CK差异不显著. 叶片相对含水量随着土壤相对含水量的升高而升高,CK叶片的相对含水量为56.39%,SWC45和SWC60处理下的叶片相对含水量与CK差异不显著,SWC75和SWC90处理下的叶片相对含水量显著高于CK.
-
甘平叶片中氮(N)含量随着土壤含水量的增加而增加,SWC75处理下的叶片N含量显著高于其余处理;不同水分处理下叶片磷(P)含量无显著差异;叶片中CK的钾(K)含量最低,SWC45和SWC60处理的叶片K含量高于CK但差异不显著,SWC75处理的叶片K含量最高且显著高于其余处理,与CK相比高32.97%;SWC45处理的叶片钙(Ca)含量最低,与CK相比无差异,但显著低于其他处理;SWC60,SWC75和SWC90处理叶片中Ca含量显著高于CK和SWC45,其中SWC75处理Ca含量最高. 结果还表明,随着干旱胁迫的减轻,叶片对Ca的吸收增加;SWC45处理的叶片中镁(Mg)含量显著低于其余处理,SWC75处理的叶片Mg含量最高(表 9).
-
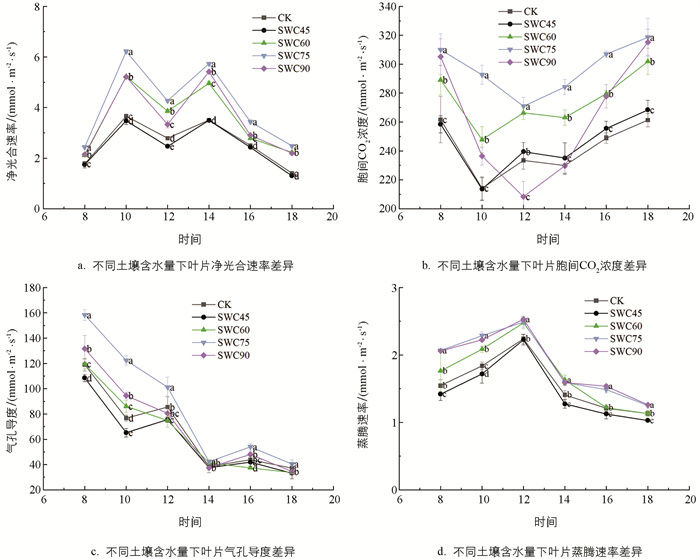
从图 3可以看出,SWC75处理下的植株光合指标显著高于其余处理,各处理的Pn日变化整体呈现“M”状,在12:00出现“午休现象”,且Pn随着土壤含水量的升高呈先升高后降低的趋势;与CK相比,SWC75处理的Pn显著提高(p<0.05).
各处理中以SWC75的Pn最高,且在全日各时段均高于其余处理;各处理的Ci日变化整体呈现“V”型,除SWC45处理外,其余处理与CK差异均达显著水平(p<0.05),且SWC75和SWC90处理的各时间段Ci显著高于CK(p<0.05),SWC90处理Ci日变化最剧烈. 各处理的Gs日变化整体呈现下凹型,在整日变化中,SWC75处理的Gs日变化显著高于其余处理(p<0.05),SWC90处理次之,处理间SWC75的Gs整日都高于其余处理. 各处理的Tr的日变化总体上呈倒“V”型,8:00-12:00各组处理Tr均不断升高,12:00后Tr开始降低. CK日均Tr为1.56 mmol/(m2·s),SWC45处理日均Tr与CK无显著差异(p>0.05),SWC60,SWC75,SWC90处理日均Tr显著高于CK(p<0.05),其中SWC75和SWC90处理Tr始终高于其余处理.
-
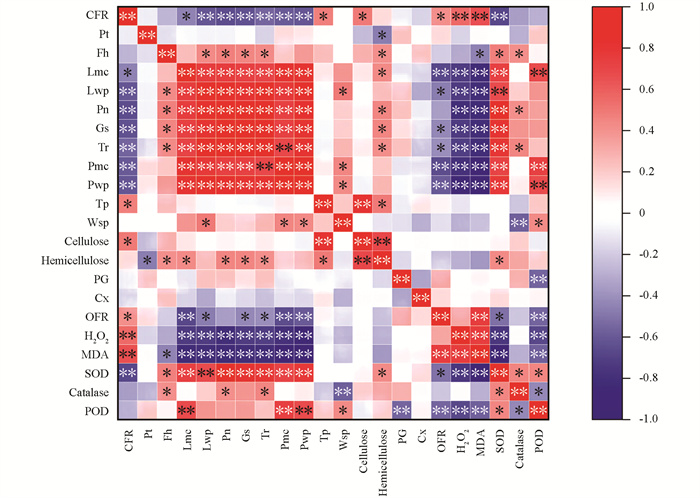
如表 10所示,根据主成分分析结果可知,叶片含水量、叶片水势、净光合速率、气孔导度、蒸腾速率、果皮含水量、果皮水势、果皮超氧阴离子摩尔质量、果皮过氧化氢摩尔质量、果皮丙二醛摩尔质量、果皮超氧化物歧化酶活性、果皮过氧化物酶活性(第一主成分中红色字体)是引起裂果的主要因素.
相关性分析可知,甘平果实裂果率与叶片水势、净光合速率、气孔导度和蒸腾速率呈极显著负相关,与叶片含水量呈显著负相关. 甘平果实果皮生理指标中,裂果率与过氧化氢摩尔质量和MDA摩尔质量呈极显著正相关,与O2-摩尔质量呈显著正相关;裂果率与果皮含水量、果皮水势、总果胶质量分数、纤维素质量分数及超氧化物歧化酶活性呈极显著负相关(图 4).
2.1. 土壤水分对甘平裂果率的影响
2.2. 正常果与裂果果实生理特征比较
2.3. 不同土壤含水量处理对果实生理的影响
2.3.1. 不同土壤水分处理下甘平果实生长动态
2.3.2. 不同土壤水分处理下果皮含水量和水势的差异
2.3.3. 不同土壤含水量下果皮细胞壁物质质量分数及水解酶活性的差异
2.3.4. 不同土壤水分处理下果皮活性氧及丙二醛(MDA)代谢的差异
2.3.5. 不同土壤水分处理下果实矿质营养的差异
2.4. 不同土壤水分处理对植株生理的影响
2.4.1. 不同土壤水分下叶片含水的差异
2.4.2. 不同土壤水分下叶片矿质营养的差异
2.4.3. 水分处理对植株光合作用的影响
2.5. 裂果因素的主成分分析
-
甘平是优良晚熟柑橘新品种,具有较高的商业价值,但是甘平高裂果性严重影响其大面积推广. 关于柑橘裂果因素前人已有不少研究报道,但是结论分歧较大[1-3, 33]. 本研究田间调研显示甘平裂果发生在盛花后90~180 d,裂果高峰集中在盛花后120~150 d,即果实快速膨大期. 甘平裂果主要是果皮纵、横向裂的外裂形式,短时间内大幅度的土壤水分变化使根系快速吸收水分,而果皮生长速率低于果肉生长速率,果实内部膨压增加,导致表皮胀裂而出现裂果. 本研究发现,75%的土壤含水量及20%以内土壤水分变化幅度能有效降低甘平的裂果,所以控制土壤含水量变化的平衡对防控裂果具有重要意义.
适宜的土壤水分会增加果皮的硬度和厚度,从而有利于果皮发育进而缓冲生长压力、减轻裂果[34]. 本试验中土壤含水量75%时,甘平果皮的厚度及硬度均最高,果皮的强度和延伸性决定了果实的抗裂性,并与果皮细胞壁物质的组成密切相关[35]. 果皮中总果胶和纤维素质量分数与甘平裂果率呈显著负相关,随着干旱胁迫程度的加重,果实中果胶和纤维素大量降解,水解酶活性增强,果皮细胞壁稳定性降低,裂果也随之增多,这与前人研究结果相一致[36-37]. 表明水分在果皮的细胞壁代谢和发育中发挥重要作用,影响果皮细胞壁的强度与完整性,进而影响裂果的易感性.
活性氧代谢的失衡会加剧细胞损伤,增强膜质过氧化水平,降低植物抗性[38],本试验主成分和相关性分析结果显示H2O2,MDA和O2-摩尔质量与甘平裂果率呈显著正相关,抗氧化酶活性与甘平裂果率呈显著负相关. 活性氧代谢失衡会增加果皮对环境变化的敏感性,进而导致果实开裂[31]. 在干旱和水分胁迫下O2-,H2O2和MDA摩尔质量均增加,造成细胞损害,导致裂果增多;在土壤含水量75%时,抗氧化酶活性高,活性氧累积量低,裂果最少. 表明水分调控可以通过维持果实代谢平衡进而减少裂果.
矿质元素是柑橘正常生命活动必不可缺的营养成分,营养物质不足会导致果实中的发育和代谢紊乱. 研究结果显示,不同土壤水分处理下,甘平中的N,K和Ca差异较大. N影响果树的生理生化过程和植株长势[39];K可以维持高渗透压和膨压,有研究表明,陷痕果果皮中的K含量高于正常果皮[40]. 在春季或果实发育早期施用钾肥可促进果皮发育,增加果皮厚度,增强果实抗开裂能力,减少收获前果实开裂[41]. Ca2+含量与柑橘裂果呈显著负相关,如红江甜橙[2]和锦橙[3]的裂果及Ca含量显著低于正常果,这与本试验的研究结果一致. 柑橘果实的生长发育和品质形成需要叶片供应养分,叶片各营养元素的丰缺对柑橘的产量和品质有重要影响[42]. 周上铃[43]报道,水分亏缺会降低柑橘叶片对N,P,K,Ca等矿质元素的吸收,进而影响果实对矿质营养的吸收与利用,本试验结果也证实了维持适宜的土壤含水量可以促进植株内营养物质的运输,减少裂果.
-
甘平裂果时间主要发生在果实膨大期,即每年7-8月,属于外裂果. 土壤相对含水量75%、变化幅度20%以内能有效控制裂果发生,该水分条件下甘平果皮细胞壁物质相对稳定,抗氧化酶活性较高,活性氧代谢平衡,植株和果实对营养物质的吸收较强,甘平裂果率达最低. 因此,在膨大期合理进行水分管理,保持稳定适宜的土壤含水量是促进甘平果实发育和植株生长、降低柑橘果实开裂的有效措施.