


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
白星花金龟(Potosia brevitarsis Lewi)隶属于鞘翅目(Coleoptera),金龟总科(Scarabaeoidea),花金龟科(Cetoniidae),星花金龟属(Protaetia)。白星花金龟一年发生1代,幼虫具有趋腐性,适应力强,在畜牧业发展较好的省份发生较为严重[1-2]。成虫可以取食为害粮食、蔬菜、水果等多达几百种寄主,尤其喜食葡萄、苹果、柑橘、玉米等作物的果实且常群集为害,能对取食部位造成酸腐现象,产生次生病害,而且成虫飞翔能力强,是农业、林业生产中的重要害虫[3-4]。目前白星花金龟的防治方法主要有3种:一是农业防治,即深耕漫灌[5]。二是基于成虫具有假死性,采用人工震落并捕捉的机械防治方式[6];利用成虫的趋糖性和趋腐性,利用糖醋液或腐烂的水果进行诱捕[7]。三是化学防治,有机磷杀虫剂、除虫菊酯类杀虫剂或植物源杀虫剂均可防治此虫[8-9]。然而白星花金龟成虫常为害成熟果实,为保证产品的安全性,不宜采用化学农药进行防治,若长期、不合理使用化学农药,容易使害虫产生抗药性,并且残留药物还会极大威胁环境及人畜健康[10],使得害虫发生与治理形成恶性循环。
昆虫病原真菌(Entomopathogenic Fungi,EPF)凭借资源丰富、杀虫广谱、致病性强、可在种群中自然流行、对环境友好、对人畜安全等特点[11],而且一些菌株结合土壤宿存能力,可以长期控制虫害发生[12],备受微生物领域学者以及植保科研工作者的青睐。目前国内外已有一些以EPF为主要成分而用于白星花金龟绿色防控的成功案例,其中以白僵菌(Beauveria sp.)[13-14]、绿僵菌(Metarhizium sp.)[15-16]较为普遍。然而,EPF作为控制自然界昆虫种群数量的主要因子,目前已经发现有750~1 000种,分属于真菌界的12个纲[17]。从现有研究来看,用于防治此类害虫的生防微生物资源仍然缺乏,因此,筛选新的EPF菌株能够丰富白星花金龟的防治资源,拓宽防治手段。本研究从一只罹病白星花金龟幼虫虫体上获得一株EPF,通过形态学鉴定和ITS-rDNA序列分析确定菌株的种类,然后初步研究不同培养基组分对该菌株生长的影响,为白星花金龟绿色防控提供备选菌株。
全文HTML
-
2019年10月,在昆明市盘龙区云南农业大学后山马铃薯试验田进行地下害虫调查时,发现白星花金龟幼虫为害取食马铃薯地下部分。将采集的幼虫带回实验室进行人工饲养。饲养过程中发现1头幼虫尸体,体表布有白色菌丝,将其保湿培养后虫体表面布满墨绿色孢子,于4 ℃条件下保存,并编号为Ma20191123。
-
马铃薯葡萄糖琼脂(PDA)培养基:将新鲜马铃薯去皮切块,然后称取200 g进行煮汁过滤,葡萄糖20 g,琼脂20 g,蒸馏水1 000 mL;萨氏葡萄糖琼脂酵母浸膏(SDAY)培养基:葡萄糖40 g,蛋白胨10 g,酵母浸粉10 g,琼脂20 g,蒸馏水1 000 mL[18]。
-
利用无菌针头轻轻挑取虫体表面的菌丝,采用点接的方式接种于SDAY培养基上,倒置于(25±1) ℃、光照条件16L∶8D的恒温培养箱中培养。3~4 d后待重新长出菌丝后,利用接种环,采用平板划线的方式将无污染菌丝接种于新的培养基上,倒置于相同的培养条件下恒温培养,直至平板上长出的菌落为同一形态。
-
挑取纯化后的Ma20191123菌株成熟分生孢子,接种于SDAY培养基中央,倒置于(25±1) ℃、光照16L∶8D的条件下培养,每个菌株设置5个重复,其中3个用于培养特征观察,2个用于形态学鉴定。每天观察菌落的形态特征及颜色。待菌落长至3~4 d后,挑取少量菌丝于载玻片上,滴加0.05% Tween-80水溶液20 μL,慢慢盖上盖玻片,置于垫有湿润滤纸的培养皿中,然后置于相同条件下培养。定期观察菌丝生长情况。培养8 d后待菌落产生分生孢子时,挑取菌落中央的分生孢子制作玻片,并在光学显微镜下观察其形态并测量大小。
-
用1 mL无菌枪头刮取少量培养基表面的新鲜分生孢子置于无菌研钵中,用液氮快速研磨成粉末状,加入到1.5 mL的离心管中。DNA提取的后续步骤参考真菌基因组DNA提取试剂盒(北京索莱宝科技有限公司),提取的DNA用分光光度计测量其浓度,OD260/OD280=1.82,之后保存在-20 ℃。
真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)由昆明硕擎生物科技有限公司合成。以提取的菌株DNA为模板进行ITS-PCR扩增反应,反应体系(25 μL):12.5 μL 2×PCRmix(北京康为世纪生物科技有限公司),1 μL ITS1,1 μL ITS4,1 μL模板DNA,9.5 μL ddH2O。扩增程序:94 ℃预变性4 min,94 ℃变性1 min,57 ℃退火1 min,72 ℃延伸1.5 min,共35个循环;72 ℃延伸8 min;4 ℃保存。PCR产物采用1.2%的琼脂糖凝胶电泳进行检测,记录凝胶成像结果,并对ITS-PCR扩增产物进行测序,由昆明硕擎生物科技有限公司测序。
将菌株的ITS序列结果去除两端质量不好的序列,最终提交至NCBI(
http://blast.ncbi.nlm.nih.gov/Blast.cgi )网站,通过BLAST比对后,筛选并下载相似性最高的典型菌株序列。运用MEGA 11软件,通过邻接法(Neighbor-Joining,NJ)构建系统发育树[19]。重复取样1 000次进行自展值(bootstrap)分析来评估系统进化树的拓扑结构的稳定性。 -
采用点接的方法,将纯化后的菌株分别接种至PDA和SDAY培养基中央,每个处理重复6次。然后倒置放入(25±1)℃、光照16L∶8D的条件下恒温培养。从第2天开始利用十字交叉法测量菌落横、纵直径,直至14 d。单个菌落的产孢量,利用直径为6 mm的打孔器从菌落的中心点至边缘距离的1/2处打孔截取小菌块,然后置于含有10 mL、0.05 % Tween -80无菌水的三角瓶中,磁力搅拌10 min,打散孢子团,获孢子悬浮液。然后利用血球计数板计算孢子浓度,若孢子浓度过高,以10倍稀释至可数清为止。
-
试验数据利用Microsoft office 2019软件进行整理后,再用SPSS 20.0软件进行数理统计分析,采用Duncan’s新复极差法进行多重比较分析。
菌落直径(cm)=(菌落横径+菌落纵径)/2;
菌落直径日增长量(cm/d)=平均菌落直径/培养天数;
单位面积菌落含孢量(孢子/cm2)=平均每小格孢子数×400×104×稀释倍数/菌饼面积。
1.1. 供试菌株
1.2. 供试培养基
1.3. 试验方法
1.3.1. 菌株分离与纯化
1.3.2. 培养性状及形态鉴定
1.3.3. 菌株分子鉴定
1.3.4. 菌株生长速率及产孢量测定
1.4. 数据处理
-
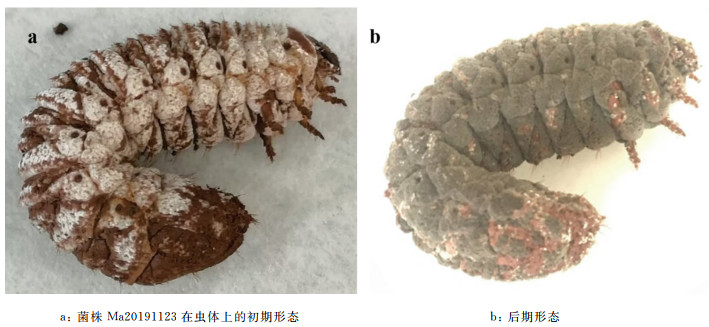
白星花金龟幼虫被Ma20191123菌株感染后,初期行动呆滞,身体呈萎靡状态。待幼虫死亡后整个虫体变僵硬,2~3 d后表面长出白色絮状菌丝,随着时间的延长菌丝布满整个虫体(图 1)。4~5 d开始出现白色分生孢子,随后2~3 d孢子成熟,整个虫体布满墨绿色孢子。从幼虫感病、病变、死亡及产孢整个过程平均为6~8 d。
-
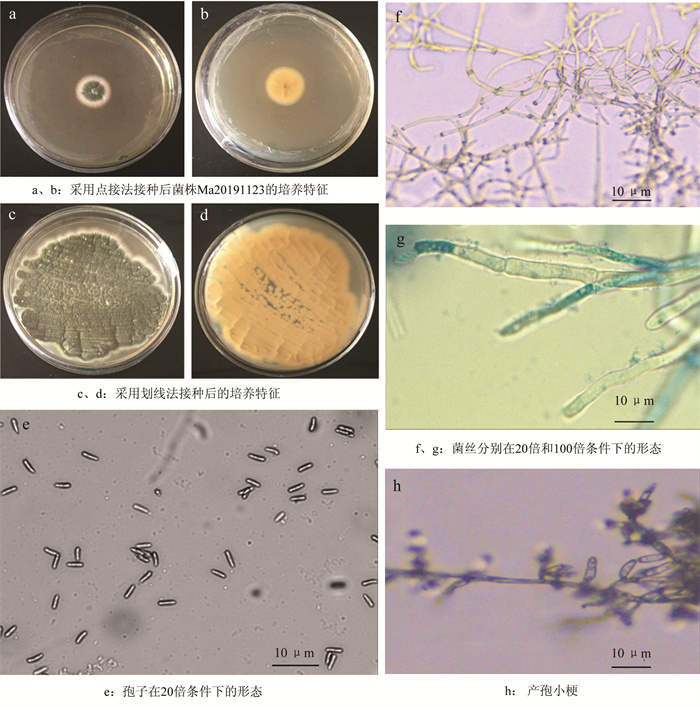
菌株Ma20191123在SDAY培养基上的菌落呈绒毛状或絮状,最初白色,中心部位形成一个菌丝密集小凸起。菌落边缘规则平整圆滑。产孢从中心位置开始,初期形成浅绿色分生孢子堆,随着培养时间的延长,菌落逐渐扩大,绿色面积变大且颜色加深,孢子由浅黄绿色变为墨绿色,菌落周围可见分散型的白色菌丝(图 2a、c)。培养基背面呈橙黄色(图 2b、d)。
菌丝具分枝有隔、透明、光滑,直径1.46~2.17 μm(图 2f、g)。分生孢子梗上多为2~3个瓶梗,瓶梗柱形,(5.1~13.8) μm×(1.7~3.5) μm(图 2h)。分生孢子为无色单细胞,柱状,链状整齐排列,大小为(5.51~6.49) μm×(1.80~2.17) μm(图 2e)。菌落、产孢细胞和分生孢子等形态特征与蒲蛰龙和李增智(1996)描述的罗伯茨绿僵菌基本一致[18]。
-
经测序后,获得一段498 bp长的片段。经BLAST比对后,该菌株的序列与数据库中大部分罗伯茨绿僵菌的相似性均能达到96%以上。为进一步明确该菌株与几种常见绿僵菌的亲缘关系,分别选择已公布的罗伯茨绿僵菌、金龟子绿僵菌(M. anisopliae)、平沙绿僵菌(M. pingshaense)、贵州绿僵菌(M. guizhouense)、棕色绿僵菌(M. brunneum)和黄绿绿僵菌(M. flavoviride)等的ITS序列构建系统发育树。结果发现,Ma20191123菌株序列以较高的自展值与罗伯茨绿僵菌聚在进化树的同一个分枝上,与其他种明显分开。序列比对结果与形态学鉴定结果相符。
因此,结合形态学鉴定结果、菌株ITS序列与数据库中的相似性比对结果以及基于ITS序列的同源性分析,最终确定Ma20191123菌株为罗伯茨绿僵菌。
-
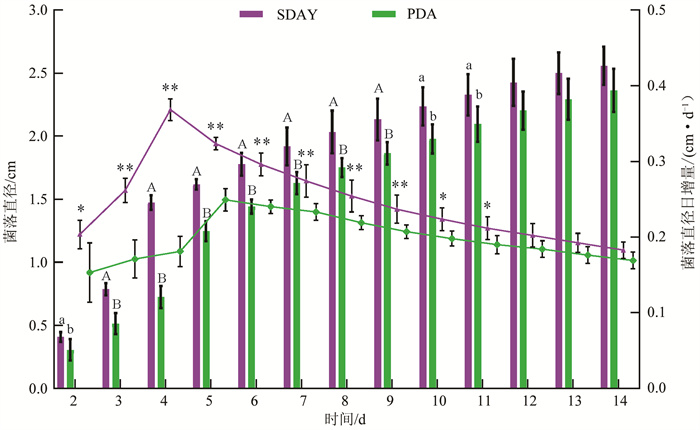
Ma20191123菌株在SDAY和PDA等2种培养基上均可以生长(图 4)。从整个培养过程来看,菌株在SDAY培养基上的长势优于PDA培养基:在SDAY培养基上,菌落直径从第2天的(0.41±0.04) cm扩大到第14天的(2.56±0.15) cm;在PDA培养基上,菌落直径在相同时段内从(0.31±0.09) cm扩大到(2.36±0.17) cm。从菌落直径日增量来看,利用SDAY培养基,2~4 d内菌落生长迅速,且第4天达到生长高峰(0.37±0.01) cm/d;相比而言,利用PDA培养基,菌落在2~5 d内生长迅速,第5天为生长高峰期(0.25±0.02) cm/d。随后菌落在两种培养基上的菌落直径日增量均随培养时间的延长呈下降趋势,直到第14天,菌落直径日增量分别降至(0.18±0.01) cm/d和(0.17±0.01) cm/d。
从产孢效果(初始产孢时间、第14天后的产孢量)来看(表 1),该菌株在PDA和SDAY培养基上分别从第5天和第6天开始产孢;第14天后的产孢量分别为(1.24±0.47)×107、(1.66±0.25)×107孢子/cm2,两者差异不显著(F1,10=3.11,p=0.11)。
2.1. 菌株种类鉴定
2.1.1. 白星花金龟幼虫被Ma20191123菌株侵染后的感病特征
2.1.2. 菌株形态特征
2.1.3. 系统发育分析
2.2. 罗伯茨绿僵菌Ma20191123在不同培养基上的生长情况
-
EPF是一类寄生于昆虫体表和体内的真菌,而且其对环境友好,杀虫方式独特,且不易使害虫产生抗药性,在农林业害虫绿色防控中具明显优势[20-21]。本研究从自然感染条件下的白星花金龟幼虫体表分离获得一株EPT,通过菌落颜色、形态学特征,初步确定Ma20191123菌株为罗伯茨绿僵菌。在此基础上,通过测序Blast比对和构建系统发育树,证实了该菌株为罗伯茨绿僵菌,将形态学和分子生物学结合[22-23],使得鉴定结果更为准确可靠。
不同EPF对培养基的营养要求和其对化学物质的敏感度是不同的,同时,培养基的营养组分及其比例也影响菌种的生长和发育以及杀虫效果[24-25]。碳作为生物合成的骨架,可以利用葡萄糖、蔗糖、马铃薯淀粉、甘油、乳糖、麦芽糖、D-果糖、D-山梨糖、D-甘露糖、i-肌醇、D-海藻糖等[26-28];氮素主要用于合成氨基酸、蛋白质、核酸和细胞质等主要成分,可以利用无机氮如(NH4)2SO4、NH4NO3、KNO3、NaNO3,有机氮如蛋白胨、花生饼粉、酵母膏、玉米粉、黄豆粉、鱼粉、麦麸等,以及谷氨酸、色氨酸和组氨酸等其他氮源的作用[29-30];矿物质元素的功能主要表现在作为菌株细胞构成的主要成分、酶的组成部分、维持酶的活性,调节细胞的渗透压、氢离子浓度以及氧化还原电位等,一般包括硫、磷、镁、钾、钠、钙、铁等[18];铜、锰、锌等微量元素还可以刺激菌株的生命活动[31]。目前PDA和SDAY培养基是用来分离真菌的常用培养基[32-34],本研究将罗伯茨绿僵菌Ma20191123转接到上述2种培养基上,该菌株在PDA培养基上除产孢时间较早外,生长速率、产孢量均以SDAY培养基最好,此结果与张松影等(2011)分离出的黄绿绿僵菌Mf82菌株在SDAY培养基上的长势具有相似的结果(菌落增长量2.59 mm/d、产孢量17.34×107孢子/cm2)[35];而刘思雨等(2017)从罹病暗黑鳃金龟幼虫上分离得到的黄绿绿僵菌Ma130821菌株在PDA培养基上生长最快(菌落直径日增量:0.21 cm/d),产孢时序最早(第6天开始产孢)、产孢量最大(1.49×106孢子/cm2)[36];李佳颖等(2013) 研究发现分离自华南地区土壤的黄绿绿僵菌SM076菌株在淀粉作为碳源的PDA培养基上生长最快,15 d的菌落直径为60.2 mm、产孢量为5.647×108孢子/平板 [37]。造成这种差异的原因可能是EPF存在的地理来源特异性和寄主专化性,致使不同地理来源、不同寄主来源的绿僵菌菌株在菌落形态、生长速率、产孢量等生物学特性以及致病力等方面存在很大差异[38-39],但具体原因还有待于进一步研究。
除了受营养物质影响之外,外界环境因素的变量对EPF的生长发育也有重要影响,只有在适宜的环境条件范畴下才能促使菌落正常地生长和发育[40-41]。目前国内外有不少学者针对温度、湿度、pH值和光照等对绿僵菌生长的影响进行研究,确定绿僵菌生长的最适温度范围在25~27 ℃[42-44],最适湿度范围在>85%[45-46],最适酸碱范围在6~7[29, 47],最适光照条件为交替光照[48-49]。本研究分离得到的罗伯茨绿僵菌Ma20191123对培养条件的要求不是很苛刻,在(25±1) ℃、光照16 L∶8 D、相对湿度80%的条件下即可正常生长产孢。但是该菌株培养条件研究是在室内恒定条件下进行的,不同于田间环境因子是复杂且随时变动的,往往造成EPF的田间防治效果不稳定;而且从EPF接触害虫体壁、孢子萌发、侵入昆虫体壁、体内繁殖,到引起昆虫死亡,每个过程均需要一定时间,致使在实际防虫应用中存在明显的时滞效应[50]。化学防治的高效性与速效性[51-52]正好弥补了EPF生防的缺陷,但大量且频繁使用化学农药引发了一系列负面影响[53]。因此,为弥补双方的不足,即解决EPF致死缓慢问题和减少化学农药用量、降低害虫抗药性风险、保护环境,最有效的途径就是将EPF与化学农药联合使用。首先,在化学农药的胁迫下昆虫可能会减少其对体表的清理行为,从而增加EPF孢子附着在昆虫体表的概率[54];其次,化学农药会降低昆虫的取食率,使昆虫更容易处于饥饿状态,从而降低其防御EPF的侵染能力[55],同时化学农药还会降低昆虫的免疫力及对EPF的抵抗力[56]。很多学者长期以来都在致力寻找能够与EPF配伍施用的化学农药。例如,常静等(2015)利用3种杀虫剂与绿僵菌混用后发现:茚虫威与绿僵菌混用增效作用最强,沙葱萤叶甲3龄幼虫的LT50较单施绿僵菌缩短了8.12 d;阿维菌素与绿僵菌混用后LT50缩短了7.60 d;鱼藤酮与绿僵菌混用后LT50缩短了6.45 d[57]。除此之外,将卵孢白僵菌NEAU30503与陶丝本(3000×)混用防治蛴螬[58],球孢白僵菌YB8分别与辛硫磷、噻虫嗪、噻虫胺、高效氯氟氰菊酯和高效氯氰菊酯联合使用防治韭菜迟眼蕈蚊[59],以及金龟子绿僵菌CQMa421与吡蚜酮联用防治白背飞虱[60]等均能起到协同增效作用。然而并非所有化学农药与生防菌混用时均起到增效作用。例如,绿僵菌MBJ2-2与多菌灵、甲基硫菌灵、代森锰锌混用后,在高、中、低浓度下均完全抑制孢子萌发、菌丝生长与产孢[61];球孢白僵菌HFW-05与60%唑醚代森联配伍施用后对小菜蛾2龄幼虫的校正死亡率降低了49.9%[62]。由此可见,EPF不同菌株对化学农药的敏感性均存在差异,不能盲目混用,其相容性研究是保证菌株发挥高致病性、化学农药发挥速效性的前提[63],选择相容性好的低毒化学农药与EPF配合使用,在提高EPT杀虫速度的同时,又可以有效降低因大量使用农药而带来的诸多负面影响,对害虫绿色防控具有双重意义。因此,为提高罗伯茨绿僵菌Ma20191123的杀虫速度,测定化学农药对其孢子萌发效果、菌丝生长以及产孢量的影响是后期研究的重点内容。