





-
北盘江地处云贵高原的斜坡地带,地形起伏大,峡谷典型,为东南季风与西南季风的交汇过渡区,雨量充沛,海拔低热量条件好,生境极特殊,是“中亚热带东部山地、山原湿性常绿阔叶林地区”和“中亚热带西部高原、山原偏干性常绿阔叶林地区”的重要交错地带[1-3].由于人为干扰严重,植被退化,加上喀斯特生境渗透性强,保水能力弱,生境残酷,临时性干旱频繁,土壤瘠薄,土壤水分不足,植被覆盖率低,导致了现在的石漠化景观.
植物群落物种多样性是反映植物群落组成结构和稳定性的重要指标,是物种丰富度和分布均匀性的综合反映,能体现群落结构类型、组织水平、发展阶段、稳定程度和生境的差异[4-5].研究群落物种多样性和种间关系,可对群落的组成、结构和功能与环境的关系有一个更好的认识,为生物多样性的保护提供理论依据[6].种间联结是指群落中不同物种在空间分布上的相互关联性[7],包括空间分布关系及物种之间的功能依赖关系[8-10],描述的是物种关系外在现象,展现的是种间竞争的结果或群落的现状[11],它是群落数量和结构的重要特征之一,是群落形成、演化的基础,也是种间相互关系的一种表现形式和群落分类的依据[12],它不仅反映了现阶段群落组成的结构特点,而且在很大程度上表现了群落未来的动态、演替趋势和演替进程[13].
基于此,本研究通过对贵州北盘江流域自然群落的调查,了解其物种组成、种间联结性关系和多样性特征等,并进行综合评价,揭示和总结出该地区所处群落自然配置的基本规律,为北盘江石漠化区造林植被恢复提供一定的理论依据.
HTML
-
北盘江位于贵州北盘江属云贵高原斜坡地带,是珠江水系红水河的一大支流.北盘江流域岩石类型主要由白云岩、石灰岩、白云岩类泥质白云岩、钙质白云岩及砂页岩等组成[14],土壤主要为石灰土[15].该地区属于亚热带湿润性季风气候,为我国东南季风与西南季风交汇区,冷暖气团常交汇于此,是贵州省光照资源较为丰富的地区之一(图 1),年平均气温为18.4 ℃,热量条件丰富.研究区降雨丰富,年均降水量为1 100 mm,部分区域(晴隆至关岭一带)可达1 550 mm,但时空分布不均,夏季多暴雨[15];研究区处于“中亚热带东部山地、山原湿性常绿阔叶林地区”和“中亚热带西部高原、山原偏干性常绿阔叶林地区”的交错带,植被类型复杂.
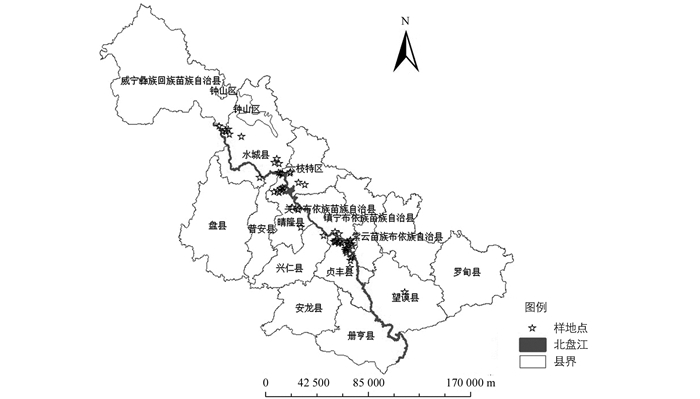
-
选取郁闭度较大,在石面、石沟和石缝生长良好的典型原始天然次生林进行样地设置,根据上中下游和海拔高度每个流域段兼顾设置典型的群落样地共78个.其中,除北盘江中段强度石漠化区(关岭至贞丰段)统一按20 m×40 m设置样地外,其余上游和下游乔木林统一按20 m×40 m,灌木、灌丛林按10 m×10 m设置样地.乔木记录树高、胸径,灌木记录地径、高度,草本记录高度、盖度等.样地内再分乔木小样方(10 m×10 m)、灌木小样方(4 m×4 m)和草木小样方(2 m×2 m)进行调查.
-
重要值计算:
乔木层[16]
灌木层[17]
草本层[18]
由于重要值考虑了频度和盖度等多个参数,因此采用重要值来测度α多样性指数[19-20],包括物种丰富度、物种多样性和物种均匀度3种指数[21].
Gleason丰富度指数:
Simpson多样性指数:
Shannon-Wiener多样性指数:
Pielou均匀度指数:
式中:S为物种数目;A为单位面积;Pi为Ii/I,Ii表示第i个种的重要值,I表示物种i所在样地各物种的重要值之和.
-
群落内种间总体关联性能有效地反映喀斯特群落植物在不同环境中的适应程度,并能充分表征植物群落的结构特征.采用零联结模型导出的方差比率法[22]定量验证植物群落的总体关联性:
式中:S为总物种数;ni为物种i出现的样方数;N为样方总数;Pi为物种出现的频度;Tj为样方j内出现的研究物种数;t为全部样方中物种的平均数;δT为所有物种出现频度的方差,ST为所有样方物种的方差,kVR为群落内物种间的总体关联指数.kVR>1,表示物种间总体表现为正关联;kVR=1,表示所有植物种间无关联;kVR<1,表示物种间总体表现为负关联.同时,采用统计量W来检验kVR偏移1的显著程度,W=kVR×N,W服从X2分布(df=N-1).若W<
$X_{0.95N}^2$ 或W>$X_{0.05N}^2$ ,则物种间总体关联显著(p<0.05);反之,若$X_{0.95N}^2$ <W<$X_{0.05N}^2$ ,则物种间总体关联不显著(p>0.05). -
根据原始数据建立2×2列联表,计算a,b,c,d值.以X2检验为基础,检验种间关联性,并建立统计量,进一步判断关联的显著性.由于取样的非连续性,采用经Yates修正过的X2统计量进行计算(表 1).
式中:N为取样总数,a表示两个物种均出现的样方数,b表示物种B出现而物种A不出现的样方数,c表示物种A出现而物种B不出现的样方数,d表示两个物种均不出现的样方数.X2<3.841(p>0.05),表示种对联结不具有统计学意义,呈独立分布;若3.841<X2<6.635(0.01<p<0.05),表示种间联结具有统计学意义;若X2>6.635(p<0.01),表示种对联结极具有统计学意义[23];当ad>bc时表示正联结,反之为负联结.
相似性指数(Ochiai指数,OI指数)的计算:
重合度指数(Dice指数,DI指数)的计算:
共同出现百分率(Jaccard指数,JI指数)的计算:
kOI,kDI,kJI这3个指数在无关联时为0,在最大关联式等于1,但不能区别出正负关联性.因此再采用联结系数kAC来进一步检验由X2统计量测出的结果可以说明种间联结程度[23],其计算公式如下:
若ad≥bc,则
若bc>ad且d≥a,则
若bc>ad且d<a,则
式中:kAC的值域为[-1, 1].kAC值越趋近1,表明种间的正联结性越强;而kAC值越趋于-1,表明种间的负联结性越强;kAC值为0,则种间完全独立.
2.1. 物种多样性分析
2.2. 种间关系分析
2.2.1. 总体关联性分析
2.2.2. 种间关联分析
-
根据统计,研究区78个样地内共调查维管植物共计445种,隶属102科288属(表 2).其中,按习性划分,乔木92种,灌木185种,藤本27种,草本118种,蕨类23种.按分类类群划分,蕨类植物9科,15属,23种;裸子植物2科,3属,4种;被子植物91科,270属,418种.其中,单子叶植物11科,42属,50种;双子叶植物80科,228属,368种.
-
科常被认为是植物分类学中较自然的演化单位,因而分析某一个区域植物区系的科是揭示该地区系特征的重要途径[24].
根据野外调查及室内鉴定,北盘江流域石漠化区维管束植物中,菊科(Compositae)包含19属,是属最多的科,其次是蔷薇科(Rosaceae)(16属),再次是禾本科(Gramineae)和蝶形花科(Papilionaceae)(14属),往下是大戟科(Euphorbiaceae)(10属).其中,含10属以上的仅有5科,占总科属的4.90%,含5~8属的科共有8科,占总科数7.84%,含2~4属的科共有40科,占总科数的39.22%,而仅有1属的为49科,占总科数的48.04%,寡属科占多数.具体统计结果见表 3.
-
北盘江流域石漠化区维管束植物中,蔷薇科含35种,是种最多的科,其次是菊科,含25种,再次是蝶形花科,含23种.往下含10种以上的分别是:禾本科(19种),大戟科(19种),桑科(Moraceae)(16种),樟科(Lauraceae)(15种),壳斗科(Fagaceae)(14种),芸香科(Rutaceae)(11种),唇形科(Labiatae)(10种).含10种以上的科共有10科,占总科数的9.80%,含5~9种的科共有19科,占总科数的18.63%,含2~4种的科为36科,占总科数的35.29%,而仅为1种的科有37科,占总科数的36.27%,单种科占多数.具体统计结果见表 4.
-
北盘江流域石漠化区维管束植物中,属内种数排第1的是悬钩子属(Rubus),所包含的种数为11种;第2是榕属(Ficus),为10种;第3是荚蒾属(Viburnum),为9种.含10种以上的属为2属,占总属数的比例有0.69%;含5~9种的属为5属,占总属数的比例为1.74%;含2~4种的属有73属,占总属数的比例为25.35%;而仅含1种的属有208属,占总属数的比例有72.22%,单种属占多数.具体统计结果见表 5.
-
北盘江石漠化区沿岸维管植物以灌木为主,共有185种,占总数的40.66%;其次为草本118种,占总数的25.93%;再次是乔木92种,占总数的20.22%;藤本和蕨类种稀少,分别为27种和23种,所占比例分别为5.93%和5.05%(表 6).从物种组成看,乔木树种以榆科(Ulmaceae)、桑科、樟科、漆树科(Anacardiaceae)、大戟科、壳斗科以及蔷薇科等为主,具有强烈的人为干扰特点;以木本植物为主的群落组成,表明石漠化生境对干扰、干旱胁迫或地貌异质性敏感,植被更新较为缓慢.
-
根据吴征镒等的世界种子植物科的分布区类型系统[25](蕨类植物通过《中国植物志》(第一卷中文版)),可将研究区植物科可分为7个分布型和4个分布变型.世界广布(1)31科,占总数的30.39%,热带性质科(2,2-2,3,4,5)79科,占总数的77.45%,温带性质科(8,8-4,8-5,8-6,14)23科,占总数的22.55%(表 7).从科的地理成分分析看,研究区植物物种组成以热带地区性质为主,与北盘江石漠化区处于亚热带与热带的气候过渡带,属湿润性的季风性气候直接相关.
288属维管植物可划分为14个分布型和8个分布变型.其中世界广布(1)28属,占总数的9.72%;热带性质属(2-7,含变型)151属,占总数的52.43%;温带性质属(8-14,含变型)105属,占总数的36.46%;中国特有属(15)4属,占总数的1.39%.热带性质占优势,属级水平的聚类相比科级水平更能体现出纬度气候相近性,体现出与气候带相匹配的植物地理成分组成特征.
属的R/T值分析,R/T为1.438 1>1,表明北盘江沿岸石漠化区维管植物属表现出明显的热带性质,与属的分布区类型分布特点相一致.详细属分布见表 8.
-
针对贵州北盘江中段典型强度石漠化区5个不同演替阶段,选择15个典型样地进行物种多样性分析,具体样地基本概况见表 9.
-
研究区样地内树种较为丰富(表 10),重要值排在前10位的主要优势树种为圆果化香树(Platycarya longipes)、复羽叶栾树(Koelreuteria bipinnata)、青檀(Pteroceltis tatarinowii)、圆叶乌桕(Sapium rotundifolium)、南酸枣(Choerospondias axillaries)、栓皮栎(Quercus variabilis)、安顺润楠(Machilus cavaleriei)、岩樟(Cinnamomum saxatile)、翅荚香槐(Cladratis platycarpa)和清香木(Pistacia weinmannifolia).其中,圆果化香树数量最多,相对密度、相对频度和相对优势度分别为0.82,0.84和0.81,占据着乔木群落中各项重要值指标的首位,在样地中占明显优势,这也与该地区以落叶树种为建群种的植被群落结构相一致.
-
总体物种Gleason丰富度指数R从大到小依次为:灌木层(4.19)、草本层(3.5)、乔木层(1.66).乔木层物种Gleason丰富度指数R较高的主要集中在乔木阶段,其次是灌乔阶段;Simpson多样性指数D和Shannon-Wiener多样性指数H差异不大;Pielou均匀度指数J以灌乔阶段略高,其余相差不大(表 11).
乔木层物种最丰富的是乔木阶段,物种越丰富,均匀度越低.这主要是由于乔木阶段的乔木优势种明显,具有较强的竞争力,占据了较多的光热条件,使得下层树种不能得到充足的光热,生长较为缓慢,而处于灌乔阶段的乔木层物种较为丰富,主要是灌乔阶段的群落还处于发育阶段,乔木层树种优势不明显,不具有较大的郁闭度,对下层树种不造成明显的遮挡,各乔木树种能接收到的光热条件相当;同时,下层树种还能够良好地生长,从而使得整体乔木树种较丰富.
灌木层物种Gleason丰富度指数较高的主要集中在灌乔阶段和灌木阶段;Simpson多样性指数较高的主要集中在灌乔阶段和灌木阶段,其次是灌乔阶段;Shannon-Wiener多样性指数较高的主要集中在灌乔阶段和灌木阶段;Pielou均匀度指数较高的主要集中在灌木阶段.但灌木层物种多样性最大的还是集中在灌乔阶段和灌木阶段,乔木阶段次之,草灌阶段总体来说物种多样性最小,这主要是因为,演替阶段不同,物种分布不同,草灌阶段还处于演替的初级阶段.
草本层物种Gleason丰富度指数较高的主要集中在草本阶段和草灌阶段,其次是乔木阶段,而Gleason丰富度指数相对较低的主要是灌木阶段.其原因可能是因为在草本和草灌阶段,没有太多的上层植被覆盖,竞争力不强,导致草本植物能够获取充足的养分,进而物种丰富度较高,多样性较丰富.而在灌木阶段,竞争力较强,草本植物和灌木生存空间相似,且较为密集和紧凑,所以灌木占据了较有利的生存空间,草本植物的生长就不乐观,所以灌木阶段的草本层物种丰富度相对较低;同理,在乔木阶段草本植物的生存空间相对灌木阶段较为空旷,不如灌木阶段紧凑,所以乔木阶段草本层植物物种相对灌木阶段就更为丰富.
-
根据方差比率法的计算,97种乔木的总体关联指数kVR为0.7912<1,因此,乔木物种总体上表现为负关联.同时,再采用统计量W来检验kVR偏移1的显著程度,W=kVR×N,W服从X2分布(df=N-1).计算出W为36.645 9>
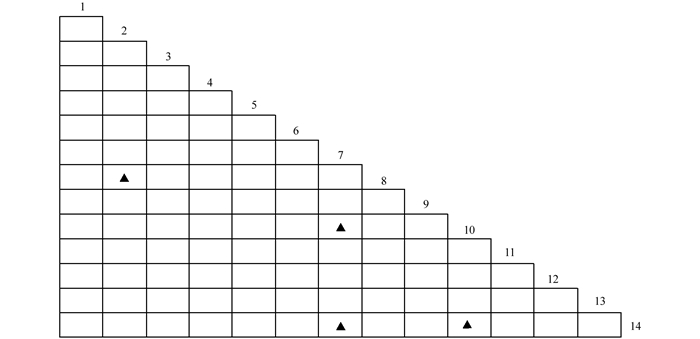
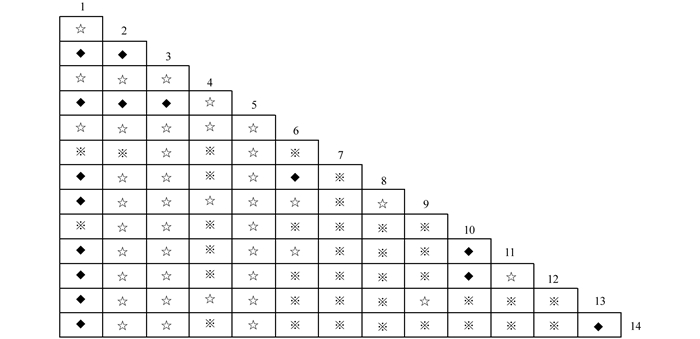
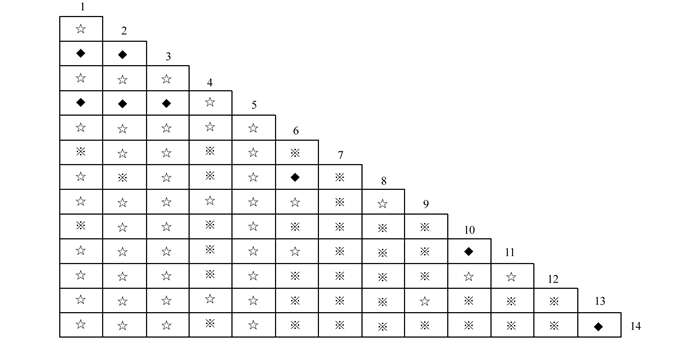
$X_{0.05N}^2$ =33.92,则物种间总体关联显著(p<0.05).根据中段典型石漠化区3个乔木阶段样地中14种乔木建群种为基础数据.计算并作图表分析其X2值(图 2)、OI指数(图 3)、DI指数(图 4)、JI指数(图 5)和联结系数(AC)(图 6).
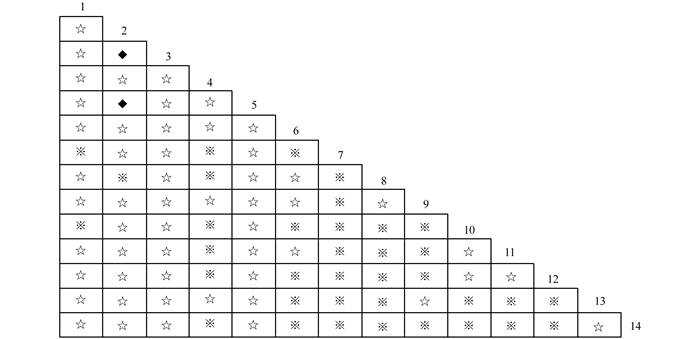
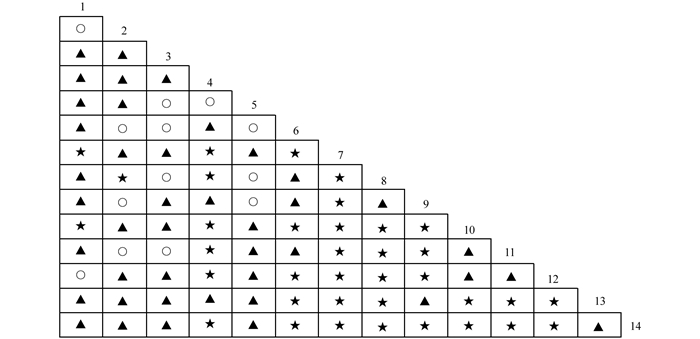
-
运用X2检验可以客观的判断树种对之间联结的显著性,对14种主要树种进行种对间X2值的计算(图 2和表 12).在所有91个种对中,显著联结的种对有4对,占总对数的4.4%,弱连结对数为87对,占总对数的95.6%.显著联结的4个种对分别是:清香木和山麻杆(Alchornea davidii),黄葛树(Ficus virens)和鸡桑(Morus australis),黄葛树和广西密花树(Rapanea kwangsiensis),鸡桑和广西密花树.
-
群落中有91个种对,OI指数,DI指数和JI指数3个指数都为0的(即在本次调查中这些种对相伴出现的几率为0.表明这些种在同一生境中同时出现的机率很小),占总对数的23.08%;3个指数大于0.5的也比较少,OI指数大于0.5的有11个种对,占总对数的比例为12.09%.DI指数大于0.5的有8个种对,占总对数比例为8.80%.JI指数大于0.5的只有2对,占总对数的比例为2.20%.3个指数都大于0.5的种对仅有2对.
根据联结系数(AC)的计算结果,在研究的91种对中,表现为正联结的种对有42对,占总对数的比例为46.15%;表现为一般负联结的有13对,占总对数的比例为14.29%;表现为强负联结的种对有32对,占总对数的比例为35.16%;显著负联结的有4对,占总对数的比例为4.40%.种间联结性总体上表现为负联结,植物种对的负联结体现了物种之间的排斥性这是长期适应不同的微环境,利用不同资源空间的结果,也是生态位分离的反应[26].
3.1. 物种组成
3.1.1. 科内属统计
3.1.2. 科内种统计
3.1.3. 属内种统计
3.1.4. 生活型统计
3.2. 区系特征分析
3.3. α物种多样性分析
3.3.1. 重要值分析
3.3.2. 多样性指数分析
3.4. 种间联结性
3.4.1. X2值
3.4.2. 联结系数(AC)
-
调查共计维管植物445种,隶属102科288属.生活型最多的是灌木.以菊科和蔷薇科占据明显优势;木本植物以悬钩子属占据明显优势,蔷薇科悬钩子属植物和菊科植物在北盘江石漠化区具较强的适应性.此外,樟科的樟属,大戟科的乌桕属,壳斗科的栲属、青冈属、栎属,芸香科的花椒属,桑科的榕属以及草本植物中黍亚科和唇形科等也同样具有较强的适应性.乔木树种的总体关联性为负关联,植被分布总体上呈现热带性质.研究区现有植被不具明显的分层现象,群落结构不稳定.
植物群落的乔木层、灌木层和草本层的Gleason物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数变化趋势基本一致,除乔木Pielou均匀度指数从大到小依次为:乔木层、灌木层、草本层,Simpson多样性指数及Shannon-Wiener多样性指数从大到小依次都为:灌木层、草本层、乔木层.
本研究区植物数量和物种丰富度与贵州茂兰喀斯特原生森林群落[26]、广西木论喀斯特森林群落[27]和广西弄岗喀斯特季雨林植物群落[28]等未发生石漠化的喀斯特区相比差异不大,与非石漠化地区相比,北盘江中段石漠化地区现有生存较好的物种数量并不少,表明北盘江中段石漠化恶劣生境下同样存在大量适应性较好的物种.
种间联结性能够反映群落的稳定状况[29],一般来讲,群落稳定性越强,种间联结越趋向于正联结;反之,群落越不稳定,群落间的排斥性就越大,种间联结越趋向于负相关[30].本研究样地中乔木树种的总体关联性为负关联,在91个种对中,负联结种对数过半.表明了群落仍处于不稳定阶段,这也可能是由于这些种生态位重叠大,具有相同的资源利用方式,种间竞争较强导致排斥性较强.
综上所述,基于北盘江流域石漠化区现有植被的样地调查,表明该区群落处于演替的初级阶段,物种相对较丰富,植物区系主要以泛热带分布属占优,植物区系的过渡性特征明显.
 DownLoad:
DownLoad: