





-
开放科学(资源服务)标志码(OSID):

-
由柑橘衰退病毒(Citrus tristeza virus,CTV)引起的柑橘衰退病对全世界的柑橘产业造成重大经济损失[1]. CTV属于长线形病毒属(Closterovirus),是正义单链RNA病毒,其基因组全长约为19 296个核苷酸,是目前已知的植物中最大的病毒[2-6]. CTV在田间存在株系分化现象[7],不同的CTV分离株会引起多种症状,包括木质部茎陷点症状,酸橙砧木的快速死亡或严重衰退症状,酸橙、葡萄柚和柠檬实生苗的枯黄症状[8]. CTV根据在指示植物上的症状可区分为强毒株和弱毒株.
植物病原体侵染植物后,往往引发寄主相应的发病机制,并激活植物防御反应等代谢途径[9],产生萜类、醇类、醛类等植物挥发性物质[10],萜类化合物是植物挥发物中含量最多且结构多样的次生代谢产物[11]. 关于病毒对寄主植物挥发性物质的影响越来越受到关注[12],如番茄黄化曲叶病毒(Tomato yellow leaf curl virus,TYLCV)显著降低番茄萜烯类挥发物的释放[13];豆荚斑驳病毒(Bean pod mottle virus,BPMV)和大豆花叶病毒(Soybean mosaic virus,SMV)侵染寄主植物后,其寄主的单萜、倍半萜以及总挥发物的释放量均显著降低[14];南方水稻黑条矮缩病毒(Southern rice black-streaked dwarf virus,SRBSDV)会诱导水稻产生特有的十三醛[15]. 目前,关于CTV侵染寄主后引发挥发性物质的变化报道极少. 为明确CTV对柑橘挥发性物质的影响,本研究采用顶空固相微萃取法(Headspace Solid-phase Micro-Extractions,HS-SPEM)结合气相色谱-质谱联用(Gas Chromatography-Mass Spectrometry,GC-MS)技术,比较并分析感染CTV茎陷点型强毒分离株CT14甜橙和健康甜橙中挥发性成分的变化,进一步应用实时荧光定量RT-PCR(RT-qPCR)技术比较分析4个萜类合成相关基因的表达差异.
HTML
-
以感染CTV分离株CT14的2年生西蒙斯甜橙实生苗嫩叶作为试验材料,以2年生西蒙斯甜橙健康实生苗嫩叶作为对照,这些材料均由西南大学柑桔研究所国家柑橘苗木脱毒中心提供.
-
分别从长势基本一致的5株健康甜橙和5株感染CTV强毒分离株CT14的甜橙嫩梢上随机采取5份嫩叶片,每份嫩叶取20 g,然后迅速加液氮研磨为粉末,准确称量2.00 g,置于20 mL螺口样品瓶中,加入2 mL饱和氯化钠溶液,并加入2 μL环己酮作为内标物,最后用聚四氟乙烯隔垫密封瓶盖旋紧后,待上机检测. 试验重复3次.
采用HS-SPME法,以二乙烯基苯/碳分子筛/聚二甲基硅氧烷(DVB/CAR/PDMS 50/30 μm)萃取头(美国Supelco公司)收集健康和感染CT14的西蒙斯甜橙叶片的挥发性物质,将样品置于50 ℃平衡20 min,顶空吸附40 min,解吸5 min.
-
参照何朝飞等[16]的方法,利用7890A/5975C气相色谱-单四极杆质谱仪(Gas Chromatography-Mass Spectrometry,GC-MS,美国安捷伦公司)对柑橘叶片挥发性物质进行分子鉴定. 色谱条件:柱起始温度为35 ℃,保持5 min,然后以3 ℃/min升至180 ℃,保持2 min,再以5 ℃/min升至250 ℃,保持2 min;采用Combi PAL气相色谱多功能自动进样器(瑞士CTC公司)不分流进样,进样口温度250 ℃,载气为氦气(纯度大于99.999%),流速1 mL/min. 质谱条件:离子化方式是电子电离(Electron Ionization,EI),电子能量70 eV,传输线温度280 ℃,离子源温度230 ℃,四极杆温度150 ℃,质量扫描范围m/z 35-450.
以C5~C25正构烷烃(色谱纯,德国Dr. Ehrenstorfer Gmb H公司)作为标准,通过其保留时间的不同计算样品中检测出化合物的保留指数(Retention Indices,RI)[17];经NIST2008和Flavour2.0图谱库检索结果,并结合相关文献共同定性[18],确定挥发性化合物的成分. 最终采用峰面积归一化法计算各成分相对质量分数进而对比内标物峰面积进行半定量,计算公式为:各成分相对质量分数(μg/g)=各成分峰面积×内标物质量(μg)/(内标物峰面积×样品质量(g)).
-
甜橙叶片总RNA用Trizol试剂盒(Invitrogen公司)提取,以RNA为模板,采用反转录试剂盒(TaKaRa公司)合成cDNA.
-
依据本研究挥发性物质分析结果与本研究室前期转录组数据,选取4个萜类合成相关基因进行表达量分析,这4个基因分别为法呢基焦磷酸合酶(FPPS)、香叶基香叶基二磷酸合酶(GGPPS)、γ-萜品烯合酶(TPIS)、(R)-柠檬烯合酶(LMS),这些基因序列均从华中农业大学柑橘基因库(
http://citrus.hzau.edu.cn/index.php )下载,用Oligo 7软件设计引物,引物序列详见表 1.使用iCycleriQ实时荧光定量PCR仪(美国伯乐公司)进行RT-qPCR. 以反转录产物为模板,用实时荧光定量PCR试剂盒(TaKaRa)进行扩增. 反应条件如下:95 ℃预变性30 s;然后95 ℃变性5 s,58 ℃退火、延伸30 s,进行40个循环;扩增后采集熔解曲线程序:95 ℃变性30 s后,从58 ℃开始,每升高0.5 ℃停留15 s采集荧光信号,直到温度升高到95 ℃熔解曲线采集完成. 为了更好消除样本之间的差异进而归一化处理实时定量PCR数据,本研究同时以甜橙Actin和GAPDH(甘油醛-3-磷酸脱氢酶)2个基因作为内参基因,内参基因引物序列分别参考贾瑞瑞等[19]和胡燕[20]的文献.
-
以Excel计算样品挥发性物质质量分数和萜类相关基因表达量的平均数、标准误,并进行t检验检测健康甜橙和感染CTV甜橙之间挥发性成分和萜类相关基因表达的差异统计学分析.
1.1. 供试材料
1.2. 挥发性物质收集
1.3. 挥发性物质的定性和定量分析
1.4. RNA提取及cDNA的合成
1.5. 实时荧光定量PCR
1.6. 数据统计与分析
-
应用SPME-GC-MS联用技术,分别对感染CTV强毒分离株CT14的甜橙和健康甜橙(对照)的挥发性成分进行分析,结果表明,与对照(健康甜橙)中检测出的59种挥发性物质相比,CTV侵染的甜橙存在55种挥发性物质,其中51种为共有,萜类化合物居多,占39种;CTV侵染后其相对质量分数显著下降者为26种,非萜类为12种,CTV侵染后其相对质量分数显著下降者为7种,共有33种物质显著下降;4种(百里香酚、香叶酸甲酯、石竹素和2-亚甲基-莰烷)为CTV侵染甜橙所特有,其中2种(百里香酚和香叶酸甲酯)为萜类化合物;8种为健康甜橙所特有,分别为α-石竹烯、β-瑟林烯、γ-芹子烯、叶醇、顺式-茉莉酮、苯乙醛、2,4-二甲基苯乙烯、4-乙基-3-壬烯-5-炔,其中3种为萜类化合物(α-石竹烯、β-瑟林烯、γ-芹子烯)(表 2). CTV侵染的甜橙所含的特有成分有些具有防御免疫作用,如百里香酚是一种单萜酚类植物精油,具有抗菌、驱虫等生物活性[21-25],石竹素具有抗炎、抗真菌和其他活性等作用[26]. 这些化合物在CTV侵染甜橙中的出现,可能与病毒侵染所诱导寄主植物产生一系列防御反应有关;另一方面,健康甜橙中所含的一些特有物质具有抗炎、抗菌的特性,而在CTV侵染后的甜橙中这些物质就消失了,如α-石竹烯和β-瑟林烯[24-25]. 由此推测,CTV的侵染可能改变了寄主的一些防御途径,使某些防御物质产生了差异.
-
与健康甜橙每克鲜叶片所含的挥发物12 255.33 μg/g相比,CTV侵染甜橙后每克鲜叶片所含挥发物的总量显著降低(8 701.69 μg/g),仅为健康甜橙的71.00%. 与健康甜橙相比,感病甜橙中有41种挥发性化合物释放量呈显著下降或未检测出. 在这些挥发性物质中,主要成分为芳樟醇、柠檬醛、柠檬烯、橙花醛、桧烯、3-蒈烯、反式-罗勒烯、香茅醛、α-松油醇、月桂烯、萜品油烯,其相对质量分数在CTV侵染甜橙和健康甜橙中均大于200 μg/g,但这些物质在感染CTV的甜橙中显著减少(表 2). 其中,芳樟醇、柠檬醛和柠檬烯的相对质量分数均高于1 000 μg/g,在CTV侵染的甜橙中芳樟醇质量分数最高,为1 267.94 μg/g;在健康甜橙中柠檬醛质量分数最为丰富,为1 843.73 μg/g. 总之,相比于健康甜橙,被CTV侵染后的甜橙中挥发物释放的总量显著降低,并且挥发成分总量的降低并不是由于个别挥发物降低或缺失导致的,而是因绝大部分挥发物都受到不同程度的抑制造成的.
-
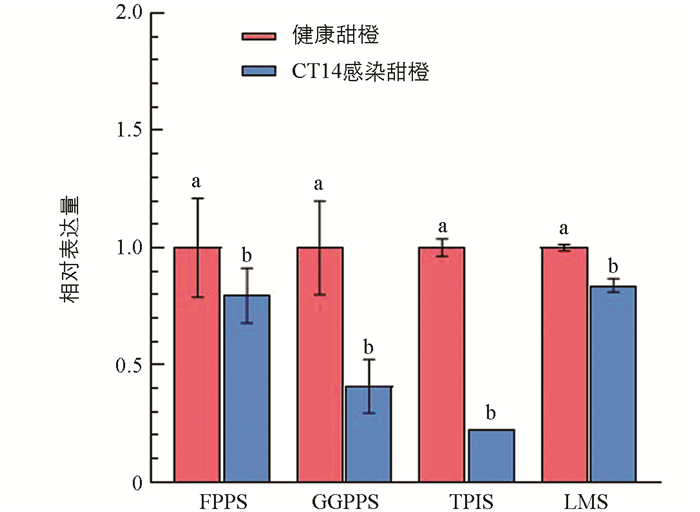
为了探索萜类成分质量分数显著减少是否与这些物质的合成途径受阻有关,依据本研究挥发性物质分析结果与本研究室前期转录组数据,本研究选择了萜类合成途径中4个关键酶基因进行表达量分析. 香叶基香叶基二磷酸合酶(GGPPS)和法呢基焦磷酸合成酶(FPPS)是萜类碳骨架合成途径中的关键合成酶,γ-萜品烯合成酶(TPIS)和(R)-柠檬烯合成酶(LMS)则分别合成γ-萜品烯和柠檬烯. 结果表明,4个萜类合成关键基因的表达量在感染CT14的甜橙中均比在健康甜橙中显著降低(图 1),这与γ-萜品烯和柠檬烯在CTV侵染后的质量分数(180.95 μg/g和1 120.22 μg/g)比在健康甜橙中(300.14 μg/g和1 565.82 μg/g)显著降低的结果相一致,暗示了萜类合成途径中关键基因表达量下调是CTV侵染后萜类化合物质量分数减少的重要原因.
2.1. CTV侵染甜橙后挥发性成分的种类比较
2.2. CTV侵染甜橙后挥发性成分相对质量分数
2.3. 萜类合成相关基因的相对表达量
-
病毒侵染寄主植物可以引起其营养成分和次生代谢等发生变化,挥发性化合物作为具有防御作用的次生代谢产物,在各种植物中广泛存在. 为了明确CTV侵染对甜橙挥发性物质的影响,本研究采用HS-SPME结合GC-MS技术对甜橙实生苗叶片挥发性成分进行分析. HS-SPME是集采样、萃取、富集及进样于一体的样品前处理技术,重现性良好,已成功应用于挥发性成分测定[27]. 研究结果发现,CTV侵染甜橙幼叶的挥发性物质总量比健康甜橙显著降低了29.00%,种类也比健康甜橙(59种)减少了4种,其中,51种为共有,26种的相对质量分数显著下降,4种为CTV侵染后所特有,8种为健康甜橙中所特有. 在这些挥发物中,萜类化合物居多,以单烯萜类、单萜醛类和单萜醇类等为主;月桂烯、3-蒈烯、罗勒烯等12种单萜烯及4种单萜醛和5种单萜醇的相对质量分数在CTV侵染甜橙后均显著降低. 这与植物病毒TYLCV,BPMV和SMV导致寄主降低植物挥发物释放量的报道一致. Cheung等[28]通过TwisterTM吸附系统及全二维气相色谱-飞行时间质谱(TD-GC×GC-TOFMS)技术发现感染CTV的田间柑橘树中月桂烯、3-蒈烯、罗勒烯等挥发性物质的相对含量均显著降低,并且提出将这些物质作为柑橘感染CTV的标志物质. 本研究检出的4种CTV侵染甜橙的特有物质分别为百里香酚、香叶酸甲酯、石竹素和2-亚甲基-莰烷. 百里香酚是一种单萜酚类植物精油,具有抗菌、驱虫等生物活性[21-25],石竹素具有抗炎、抗真菌和其他活性等作用[26],香叶酸甲酯和2-亚甲基-莰烷的功能目前尚未见报道,这些化合物在CTV侵染甜橙中的出现,可能与病毒侵染所诱导寄主植物产生一系列防御反应有关,这些物质是否也可以作为CTV的标志物质尚待进一步研究.
挥发性物质对柑橘果实感官质量有着重要作用,柠檬烯、月桂烯和柠檬醛等挥发物是柑橘的主要香气成分[29-31],并且橙花醛和柠檬醛的相互作用几乎确定了甜橙的香气[32]. 在本研究中健康甜橙和感染CTV甜橙叶片挥发性物质主要成分都是柠檬醛、柠檬烯、芳樟醇和橙花醛等,CTV的侵染显著降低了这4种挥发物的释放量,可能是造成感病柑橘果实品质降低的原因之一. 另外,柑橘挥发性物质具有较强的抗菌和抗氧化等生理活性[33],例如柠檬烯具有抗菌抑菌等作用[34-36]. Hijaz等[37]发现甜橙被感染细菌性病原-柑橘黄龙病菌(Candidatus Liberibacter asiaticus,CLas)后其释放的柠檬烯、十一烷醛的量在增加,而在本研究中CTV的侵染显著降低了甜橙叶片中柠檬烯的释放量. 因此,不同的病原引起寄主植物挥发性物质的变化有所不同.
Luan等[12]发现在中国番茄黄化曲叶病毒侵染番茄后,萜类合成途径相关基因的表达水平发生改变. 本研究选择了4个萜类化合物合成途径中的关键基因,分析了其表达是否与CTV侵染甜橙的萜类物质的显著减少一致,这4个基因分别是香叶基香叶基二磷酸合酶(GGPPS)、法呢基焦磷酸合酶(FPPS)、γ-萜品烯合酶(TPIS)和(R)-柠檬烯合酶(LMS). 在萜类碳骨架合成途径中,香叶基香叶基二磷酸合酶(GGPPS)和法呢基焦磷酸合酶(FPPS)是2个关键的合成酶,γ-萜品烯合成酶(TPIS)和(R)-柠檬烯合成酶(LMS)分别是γ-萜品烯和柠檬烯合成酶. 结果表明,这4个基因的表达量在CTV侵染甜橙后均显著下调,与其生成的直接产物γ-萜品烯和柠檬烯质量分数显著下降一致,表明CTV影响了寄主萜类合成途径进而影响了挥发性物质的释放.
-
本研究采用HS-SPEM结合GC-MS技术分析了感染CTV甜橙和健康甜橙的挥发性成分,结果表明,CTV侵染甜橙后挥发性成分总量显著降低了29.00%,且以萜类化合物为主,感染CTV甜橙中4个萜类合成关键基因表达量显著下降,与萜类挥发性物质减少一致,说明CTV侵染甜橙影响了寄主萜类合成途径和挥发性物质的释放.
 DownLoad:
DownLoad: