





-
开放科学(资源服务)标识码(OSID):

-
菜豆(Phaseolus vulgaris L.)属豆科(Leguminosae)蝶形花亚科(Papilionoideae)菜豆属(Phaseolus)植物,是菜豆属重要的栽培种之一,又称芸豆、四季豆[1],由墨西哥、中美洲中心和南美洲中心驯化为栽培种[2]. 中国有悠久的菜豆栽培历史,主要分布在黑龙江、内蒙古、吉林、云南、贵州、山西、陕西等地[3]. 我国菜豆产业目前存在品种单一、退化严重,以及种质资源命名混乱等问题,如“同物不同名,同名不同物”等现象普遍存在[4-5],因此,对菜豆种质的真实性进行区分整理,揭示其亲缘关系,对菜豆种质资源的保护与利用有重要意义.
简单重复序列(Simple Sequence Repeat,SSR),又称微卫星DNA,是指基因组中由1~6个核苷酸组成的基本单位多次重复而构成的DNA片段,因其多态性丰富、稳定性好和具共显性而备受关注. 根据来源不同,可分为基因组SSR和表达序列标签SSR(EST-SSR),其中EST-SSR标记因开发简单、快捷和低成本而在很多作物上得到应用. 徐国辉等[6]利用10对EST-SSR核心引物,对蓝莓优良品系的父本进行了鉴定,并构建了相应的DNA指纹图谱. 杨仕美等[7]基于火龙果转录组测序数据,开发出EST-SSR标记并用于火龙果种质的亲缘关系分析. 李松等[8]利用20对SSR核心引物,对云南玉溪玉米自交系进行遗传多样性分析,为云南玉米品种推广提供了理论依据. 在菜豆上,陈其福等[9]采用SSR标记分析了14个食荚菜豆品种的亲缘关系,并建立了特异性指纹图谱. 杨义杰等[10]应用8对SSR引物分析了20份芸豆品种的遗传多样性. Shabib等[11]利用SSR分子标记技术对11份菜豆品种作遗传多样性分析,为新品种选育提供了研究依据,但尚未见菜豆EST-SSR分子标记开发利用的报道.
菜豆是贵州主要夏季蔬菜之一,也有极为丰富的种质资源,但其亲缘关系和遗传背景不清晰,极大地影响了遗传育种和知识产权保护. 本文基于菜豆转录组数据,设计EST-SSR引物,建立EST-SSR标记系统;筛选出核心引物,分析了50种贵州种质和31个省外品种间的亲缘关系,并构建其分子身份证,旨在为菜豆育种亲本的选择、品种鉴定以及品种知识产权保护提供可靠的遗传学依据.
HTML
-
本研究所用菜豆种质共81份,其中50份采自贵州各地,由贵州大学蔬菜研究院提供,剩余31个品种为市面采购. 种质名称信息见表 1.
-
将菜豆转录组测序样本送至生工生物工程(上海)股份有限公司,利用Illumina HiSeq平台进行分析测序,对上机测序得到的原始序列进行过滤,去掉含接头的短序列和低质量的短序列,得到高质量的测序数据. 使用MISA软件对基因转录本进行SSR检测,根据搜索出的SSR位点及其侧翼序列,使用Primer 3完成EST-SSR引物设计.
-
将获得的81份菜豆种子浸种2~3 d后,移栽至土壤培养2~3周,直至长出2片真叶,采摘包于锡纸,放置于-20 ℃冰箱内,用于后续DNA提取. 采用北京天根生化科技有限公司的新型植物基因组DNA提取试剂盒(离心柱型)提取所有样品DNA,操作步骤按照试剂盒说明书进行. DNA提取完成后采用1%琼脂糖凝胶电泳检测DNA的完整性,采用全波长酶标仪(Thermo Fisher Scientific,MA,USA)检测其浓度与纯度,质量合格后将其存于-20 ℃冰箱中备用.
选取种子性状差异性较大的8个样品DNA为模板,利用128对引物分别进行扩增. PCR扩增体系为10 μL:模板基因组DNA 1 μL,正、反向引物各1 μL,2×Taq PCR Master Mix 5 μL(北京天根生化科技有限公司),ddH2O补充至10 μL. SSR-PCR扩增程序:94 ℃预变性4 min;94 ℃变性30 s,最适退火温度57~60 ℃反应30 s,72 ℃反应1 min,共38个循环;72 ℃延伸10 min;扩增产物放置于4 ℃冰箱中冷藏保存.
PCR产物用2%琼脂糖凝胶电泳检测,初步筛选出有多态性的引物. 将多态性较好、重复性高、稳定性好的引物对81份菜豆样品进行PCR扩增,最后使用毛细管电泳对扩增产物进行检测分析. 毛细管电泳由Qsep100TM全自动核酸蛋白分析仪完成.
-
利用毛细管电泳方法采集条带,人工读条统计100~500 bp内的条带,记录明显条带,对多态性佳的引物重复扩增至少3次,其中绝大多数带型可重复,对不稳定谱带忽略不计. 有谱带的标记为“1”,没有谱带的标记为“0”,建立数据库. 利用POPGEN 32软件分析每对EST-SSR引物的等位基因数、有效等位基因数、Nei' s基因多样性指数及Shannon信息指数;应用PIC_CALC软件计算PIC值;用NTSYS pc 2.10e软件计算相似系数,并按UPMGA方法构建亲缘关系树状图.
-
利用核心引物扩增的带型数据,赋值“0/1”形成菜豆种质的指纹代码,通过条形码和二维码生成器(
https://www.qr-batch.com/ )生成包含各供试种质基本信息的DNA分子身份证.
1.1. 试验材料
1.2. 菜豆转录组测序及引物设计
1.3. PCR扩增及产物检测
1.4. 数据处理
1.5. DNA分子身份证构建
-
利用MISA软件对菜豆转录组测序数据85 276条非冗余序列进行分析,共挖掘出11 649个EST-SSR位点. SSR标记的重复单元碱基数有单碱基重复、二碱基重复、三碱基重复、四碱基重复、五碱基重复、六碱基重复,还有复杂的重复碱基(略),对应的标记数目分别为6 610,2 389,2 458,132,36,24. 菜豆转录组EST-SSR标记主要重复单元为1~3个碱基,其中单碱基重复位点数最多,占总量的56.7%;其次是三碱基重复,占总量的21.1%. 使用Primer 3共设计EST-SSR引物30 177对,随机合成128对EST-SSR引物,分析81份菜豆种质的遗传多态性.
-
在128对EST-SSR引物中,122对引物能扩增出条带,有效扩增比例为95%. 经重复筛选,有18对引物能稳定扩增出多态性好、重复性高的条带(表 2),部分引物筛选结果见图 1.
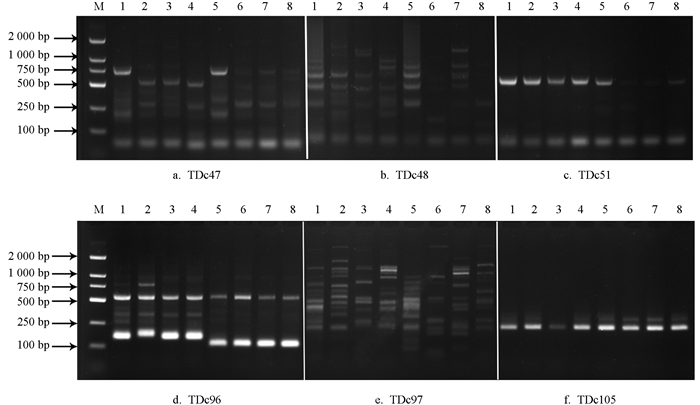
-
利用18对多态性引物对81份菜豆样品进行PCR扩增,将扩增产物进行毛细管电泳,部分种质的电泳结果见图 2. 通过POPGEN 32对统计的“0/1”数据进行分析,结果显示,18对引物在81份样品中共检测出492个多态性位点:最小多态性位点数为8个,最大为39个,平均27.3个. 等位基因数平均为2.0个,有效等位基因总数为23.0个,单对引物产生有效等位基因数为1.1~1.4个,平均为1.3个. Nei' s基因多样性指数为0.07~0.25,平均为0.18. Shannon信息指数变化范围为0.14~0.40,平均为0.30. PIC值为0.50~0.95,平均为0.88(表 3). 结果表明,筛选出的18对EST-SSR引物具有很好的多态性,81份菜豆种质也具有丰富的遗传多样性.
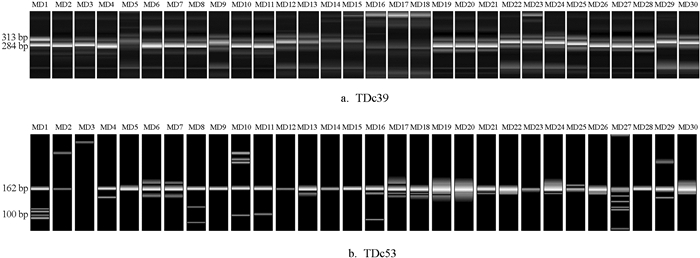
-
利用NTSYSpc 2.01e软件计算81份菜豆种质的遗传相似性系数. 聚类结果表明,遗传相似性系数变化幅度为0.15~0.65,其中MD4和MD8亲缘关系较近,相似系数为0.65,而MD37和MD77亲缘关系较远,相似系数仅为0.15. 在遗传相似系数为0.35时,可将81份种质分为3大类(图 3):第Ⅰ类包含MD1,MD3,MD4等18份种质. 第Ⅱ类中包含的种质数量最多,共计59份种质,在相似系数为0.37时,又可将其分为3个亚类,其中第一亚类包含MD24,MD25,MD30等36份种质,第二亚类包含MD56,MD58,MD63等22份种质,而第三亚类仅有种质MD57. 第Ⅲ类仅有4份来源于贵州的种质,即MD36,MD37,MD38和MD47. 省外种质(MD51~MD81)全部聚在第Ⅱ类群,极有可能与来源地有关,因为这31份品种均是来源于北方,可能其遗传背景重合度较高;而来自贵州的50份种质在3大类中均有聚类,折射出贵州种质间的遗传差异大,可以从中挖掘优异资源,在遗传育种上有较大的潜在价值.
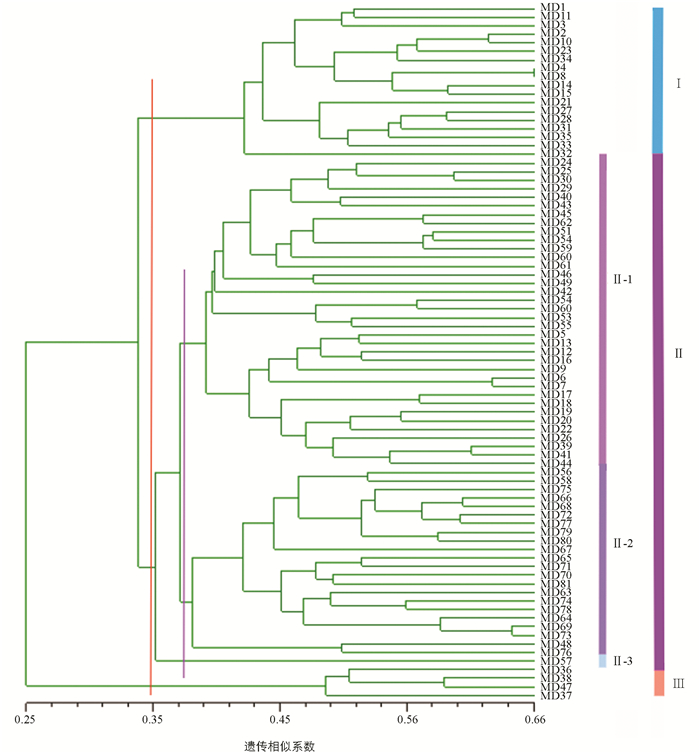
-
利用18条EST-SSR引物对81份菜豆种质进行毛细管电泳,获得每对引物扩增的带型数据,这些“0/1 ”数据可作为菜豆种质指纹图谱或分子身份证构建的直接依据. PIC值较高的引物为TDc17(0.95),TDc47(0.95),TDc66(0.94),TDc69(0.94)及TDc97(0.95)(表 3),其中TDc17,TDc66,TDc97均能鉴别出77份种质,TDc66的Shannon信息指数(0.40)最高,故选取TDc66作为核心引物,但仍有4份种质不能被识别,分别为MD42,MD49,MD57和MD63. TDc9,TDc47和TDc97等3对引物可鉴定这4份种质. 这3对引物的多态性位点数分别为33,39,37,选取多态性位点数少且具备同等鉴别力的引物对TDc9,能避免构建分子身份证的繁琐性,因此,利用引物对TDc9和TDc66扩增的EST-SSR标记,可构建全部供试种质的分子身份证,分别生成包含基本信息的条形码和二维码(表 4).
2.1. 菜豆转录组EST-SSR位点分析及引物设计
2.2. 菜豆EST-SSR引物筛选
2.3. 菜豆EST-SSR引物多态性分析
2.4. 菜豆种质的亲缘关系分析
2.5. DNA分子身份证的构建
-
相比于主观意识强,且易受环境、气候等外界因素影响的传统形态学标记,共显性强、标记数量充足、多态性高、能够高效快速鉴别种质特异性的分子标记备受关注. 菜豆作为我国主要的粮食作物,却存在种植品种单一、品种退化严重以及种质资源命名混乱等问题,因此急需开发更为可靠且方便快捷的分子标记来促进菜豆育种. 转录组数据是开发标记的理想资源[12],相较于基因组测序,转录组测序成本低、耗时短. 近年来许多果树[13]、木本植物[14]、药用植物[15]以及食用菌[16]等,均基于转录组测序开发出EST-SSR分子标记用于遗传多样性分析和品种种质鉴定及辅助育种. 本研究基于菜豆转录组数据分析结果,开发合成了128对EST-SSR引物,并验证挖掘了18对重复性好、特异性强、多态性高的引物,用于分析菜豆的遗传多样性.
根据目前的相关研究报道,当PIC>0.5时,表明引物有较高多态性;0.5≥PIC>0.2时,表明引物有中等多态性;PIC≤0.2,则表明引物多态性较低[17]. 夏春阳等[3]利用SSR分子标记分析了69份普通菜豆种质资源的遗传多样性,PIC值平均为0.659. 另外,陈琼[18]也利用33对SSR核心引物对26份菜豆DUS测试标准品种的遗传多样性进行了分析,PIC值平均为0.604. 本研究中所筛选的18对引物的PIC(0.50~0.95)≥0.5,表明这18对引物具有较高多态性,因此,本文开发的EST-SSR标记系统能用于菜豆种质的鉴定及亲缘关系分析.
-
地理来源不仅是物种亲缘关系分析的重要指标,更是影响物种遗传分化的重要因素[19]. 在长期的自交或近交下,不但会降低种群的遗传多样性,还会大大限制挖掘优良种质的可能性和培育新品种的创新性. 本研究中所用种质主要分为贵州省内及省外两类样本,省外31份菜豆样本主要来源于山东、河北和甘肃等地. 基于菜豆转录组分析结果,本研究成功完成EST-SSR分子标记的引物设计开发,筛选出18对多态性极佳的EST-SSR引物,利用这些多态性好的引物能将81份菜豆种质明确区分开,可分为3大类群. 由聚类结果可以看出,省外品种主要集中在第Ⅱ类群. 这一趋势表明这些品种的亲缘关系较近,极有可能是具有相同或相似的遗传背景,导致遗传变异水平低. 贵州省内收集的50份菜豆种质资源可被分为3大类群,且第Ⅰ,Ⅲ类群均为省内种质,仅有少量种质在第Ⅱ类群的第三亚类中,表明这50份种质遗传变异水平高,且遗传背景丰富多样. 这可能与我国西南优质菜豆种质资源交流逐年加强、少量新育成品种逐渐具备更丰富的遗传背景有关[18],因此,在杂交育种时需尽量选取来源不同的资源进行组配,以拓宽遗传背景,从而提高我国菜豆种质资源的遗传多样性[20].
-
目前,有3种构建分子身份证的方式:1) 赋值“0/1”形成字符串;2) 编码等位基因;3) 编码基因型[21]. 谢倩等[22]利用12对EST-SSR引物对59份橄榄种质进行遗传多样性分析,并构建了分子身份证. 刘伟等[23]应用SSR荧光标记法构建山东地方桃种质资源分子身份证. 陆育生等[24]利用SSR标记分析了广东黄皮种质资源的遗传多样性水平及亲缘关系,并构建了分子身份证. 本研究以“0/1”字符串生成条形码,作为构建分子身份证的直接依据;利用TDc9和TDc66作为鉴定全部供试种质的高效核心引物,以此构建81份菜豆种质的分子身份证.
本研究开发的EST-SSR标记可用于菜豆的系统发育、物种进化及亲缘关系等研究,为今后菜豆品种资源保存、利用、育种和提高育种效率等奠定理论基础,充分发挥它们在育种工作中的应用价值.
 DownLoad:
DownLoad: