





-
开放科学(资源服务)标识码(OSID):

-
细胞分裂周期相关蛋白5(CDCA5),也被称为Sororin蛋白,是一种编码人类11号染色体(11q131.1)基因的磷蛋白,其动态定位和功能受到CDK1/CyclinB和ERK2等蛋白激酶的调控[1]. 姐妹染色单体之间的内聚发生在DNA复制过程中,受到内聚蛋白的调控,并依赖于CDCA5和内聚蛋白的乙酰化[2]. WAPL是内聚的关键调控因子,它可以促进内聚蛋白从DNA中解离,CDCA5可以通过稳定内聚蛋白与DNA的结合来拮抗WAPL的作用,从而稳定姐妹染色单体的内聚. 相关研究表明,在多种肿瘤细胞周期中已经发现染色体黏连蛋白异常,黏连蛋白以及与之相关的蛋白高表达,可能是导致肿瘤发生发展的因素. CDCA5是一种潜在的生物标志物,也是多种人类恶性肿瘤的关键介质,许多细胞实验已经将CDCA5的表达上调与多种恶性肿瘤联系起来. 多项研究报道CDCA5在多种恶性肿瘤中表达上调,包括膀胱恶性肿瘤[3]、肝恶性肿瘤[4-5]、胃恶性肿瘤[6]和肺恶性肿瘤[7].
恶性肿瘤的生长是由肿瘤致癌基因或抑癌基因突变所产生的癌变细胞所驱动的[8]. 在恶性肿瘤的发生和发展过程中,许多不携带致癌突变的基质细胞类型通过与恶性肿瘤细胞相互作用而发挥功能. 免疫疗法能够改变许多晚期恶性肿瘤的治疗方式,并彻底改变恶性肿瘤的临床治疗效果. 越来越多的人认识到,恶性肿瘤微环境在肿瘤细胞发生转移的过程中发挥着至关重要的作用[9-11]. 恶性肿瘤复杂的微环境,如何适当地利用尚不清楚,因此,进一步了解肿瘤微环境中基质和免疫成分的动态调控机制,阐明肿瘤与免疫相互作用的免疫表型和验证新的免疫相关治疗靶点至关重要[12-13].
TCGA数据库收录了多种人类肿瘤(包括亚型在内的肿瘤)的临床数据,包括基因组变异、mRNA表达、miRNA表达、甲基化等,是肿瘤研究者很重要的数据来源. GTEX数据库收录了大量健康人的生物信息,该平台提供开放获取的数据,包括基因表达、QTLs及生理组织学图片等. 肿瘤免疫疗法的成功,如过继细胞转移疗法和免疫检查点抑制剂疗法,已经证明免疫细胞特别是T细胞,可以被用来消除肿瘤细胞. 作为肿瘤微环境(TME)的主要组成部分,免疫浸润物已被证明有助于肿瘤进展和免疫治疗反应,因此,更好地理解TME中的先天免疫细胞和适应性免疫细胞对于破译免疫治疗机制、定义预测性生物标志物和识别新的治疗靶点至关重要. 基于上述两个数据库信息,研究CDCA5的表达上调与肿瘤浸润免疫细胞(TIICs)及相关免疫标志物的关系,CDCA5表达与肿瘤免疫调控基因表达的关系、与抗肿瘤药物的半抑制浓度的关系,CDCA5 CNV与mRNA表达的相关性,旨在为CDCA5在泛癌中的功能作用提供新的见解,为恶性肿瘤的免疫治疗提供新的思路.
HTML
-
从TCGA和GTEX数据库下载统一和标准化的泛癌数据集[14-15],包括33种肿瘤共11 541份个体样本,10 505份癌旁组织样本. 使用R包中的rma函数(
http://www.rproject.org/https://www.rstudio.com/ )对数据进行分析,通过删除缺失和重复的结果过滤数据集,并通过log2(TPM+1)进行转换. DNA拷贝数和甲基化数据来自cBioPortal数据库(https://www.cbioportal.org/ ). -
采用Kaplan-Meier Plotter网站(
http://kmplot.com/analysis/index.php?p=background )评估患者的总生存期(OS). 采用单因素Cox回归分析评估CDCA5在预测患者的总生存期、疾病特异性生存期(DSS)、无病生存期(DFI)和无进展生存期(PFI)方面的意义. -
使用TIMER 2.0(
http://timer.cistrome.org/ )评估6种主要免疫细胞类型的浸润得分,包括B细胞、CD4+T细胞、CD8+T细胞、巨噬细胞、中性粒细胞和树突状细胞[16-17],并且评估CDCA5表达与免疫细胞浸润之间的相关性. -
通过GSCA网站(
http://bioinfo.life.hust.edu.cn/GSCA/#/ )的Drug模块分析了CDCA5表达与药物IC50之间的关系. 通过GDSC和CTRP两个药物数据库分析CDCA5表达与药物敏感性之间的关系,并绘制气泡图. 气泡的颜色代表了mRNA表达与半数抑制浓度之间的相关性[18]. -
利用人类蛋白图谱(HPA:
https://www.proteinatlas.org/ )数据库提取人类肿瘤和癌旁组织中CDCA5的蛋白水平. 利用String(https://string-db.org/ )数据库构建蛋白与蛋白相互作用的网络(PPI). 选取所有与CDCA5有交互作用的蛋白质,其中包括表达模式一致、共存、共表达、经实验验证的所有数据库蛋白质,设置最小截断值为0.900. -
首先在STRING网站(
https://string-db.org/ )上寻找与CDCA5交互的蛋白质,并设置最低置信水平为0.900,经实验验证且不超过50个的互作蛋白质;然后使用GEPIA2网站的“Slmilar Gene Detection”模块,获得TCGA数据库中所有肿瘤类型中与CDCA5相关的前20个基因,并进一步应用“correlation analysis”对CDCA5和所选基因进行成对基因Pearson相关分析;最后采用R工具“tidyr”和“ggplot2”软件包对上述数据进行KEGG通路分析[19]. -
通过GSCA网站分析不同肿瘤中CNV和甲基化与CDCA5 mRNA表达量的相关性.
-
所有数据均使用R 4.0.3软件进行分析,采用Wilcoxon秩和检验进行组间比较,两组以上比较使用Kruskal-Wallis检验. 采用Pearson进行相关性分析,以p<0.05认为差异有统计学意义. 采用R工具中ggplot 2软件包创建图形.
1.1. 数据收集
1.2. Cox回归分析和生存分析
1.3. 免疫细胞浸润分析
1.4. CDCA5表达与药物IC50相关性分析
1.5. 蛋白质水平分析
1.6. CDCA5相关基因富集分析
1.7. CDCA5 mRNA的表达量与CNV和甲基化的关系
1.8. 统计分析
-
基于TCGA数据库评估CDCA5在泛癌中的表达,结果显示CDCA5在26种肿瘤中高表达,其中24种肿瘤表达显著上调:多形成性胶质母细胞瘤(GBM),宫颈鳞癌和腺癌(CESC),肺腺癌(LUAD),结肠癌(COAD),结直肠癌(COADREAD),乳腺癌(BRCA),食管癌(ESCA),食管胃交界癌(STES),肾乳头状细胞癌(KIRP),混合肾癌(KICH+KIRC+KIRP,KIPAN),胃癌(STAD),前列腺癌(PRAD),子宫内膜癌(UCEC),头颈部鳞状细胞癌(HNSC),肾透明细胞癌(KIRC),肺鳞状细胞癌(LUSC),肝细胞癌(LIHC),甲状腺癌(THCA),直肠腺癌(READ),胰腺癌(PAAD),嗜铬细胞瘤和副神经节瘤(PCPG),膀胱尿路上皮癌(BLCA),肾嫌色细胞癌(KICH)和胆管癌(CHOL)(图 1a). 此外,本研究结合TCGA和GTEX数据库,计算了每个肿瘤中癌旁样本和肿瘤样本之间的表达差异,观察发现CDCA5在34个肿瘤中的表达显著上调,而在TGCA数据库中,其表达显著下调(图 1b).
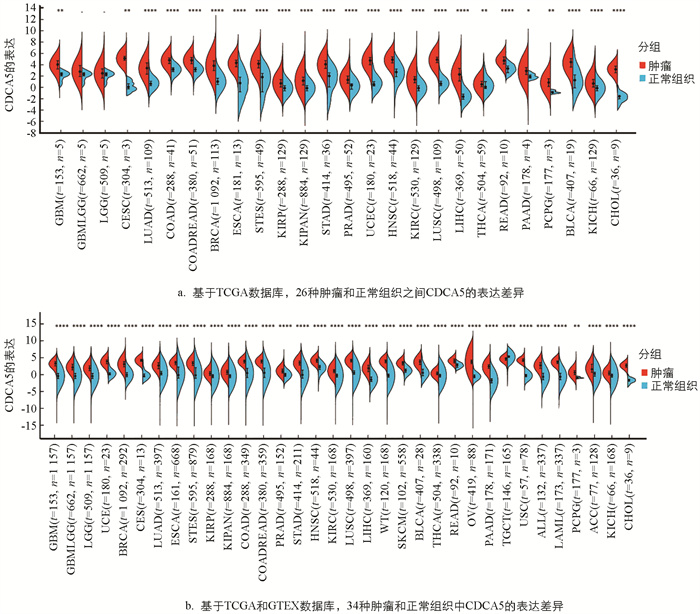
-
使用GEPIA2的“Pathological Stage Plot”模块检测CDCA5表达与各种肿瘤病理分期的相关性,发现在肾上腺皮质癌(ACC),BRCA,皮肤黑色素瘤(SKCM),LUSC,LUAD,KIRC,KICH,卵巢浆液性囊腺癌(OV),LIHC,KIRP和子宫肉瘤(UCS)中,CDCA5表达增加与肿瘤分期相关(图 2,p<0.05).
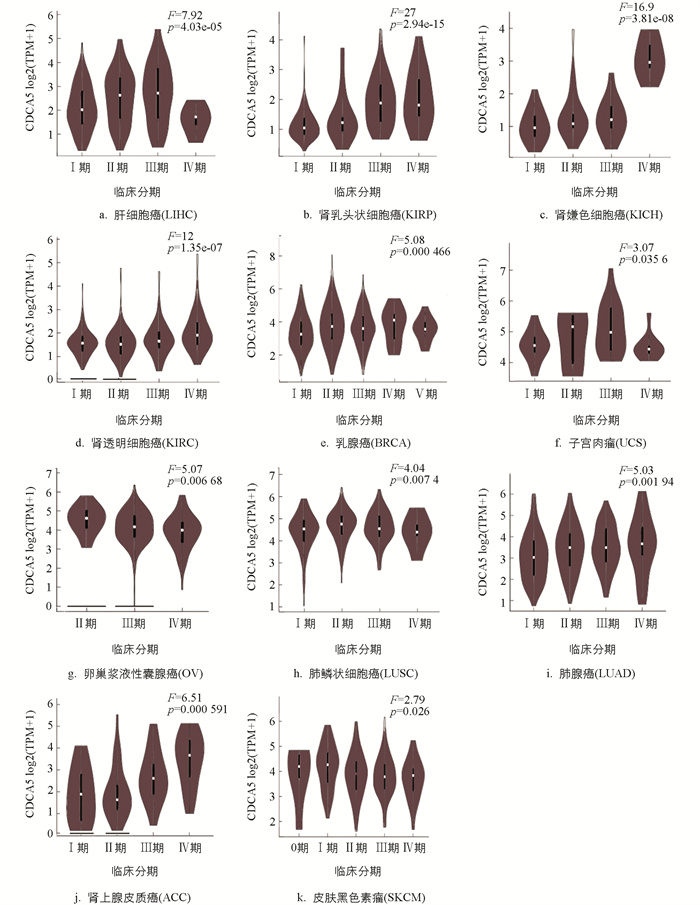
-
通过Kaplan-Meier Plotter网站探讨CDCA5表达水平与肿瘤和预后的相关性,结果显示,CDCA5的表达与肿瘤预后相关,包括乳腺癌、卵巢癌、肺癌、肝癌. 单因素Cox回归分析结果显示,CDCA5高表达是胶质瘤(GBMLGG),KIPAN,KIRP,ACC,LGG,恶性间皮瘤(MESO),KICH,LIHC,LUAD,PAAD,KIRC,PRAD,SKCM-M,SKCM和PCPG患者总生存期预后不良的危险因素,是胸腺癌(THYM)患者总生存期预后不良的保护因素. CDCA5高表达是KIPAN,GBMLGG,KIRP,PRAD,ACC,KIRC,PAAD,KICH,LIHC,MESO,PCPG,LGG,葡萄膜黑色素瘤(UVM),THCA,LUAD,BRCA和BLCA患者无进展生存期的危险因素,是卵巢癌患者无进展生存期的保护因素. CDCA5高表达是KIPAN,GBMLGG,KIRP,ACC,KICH,KIRC,LGG,MESO,PAAD,LIHC,LUAD,PRAD,BRCA,SKCM,SKCM-M和PCPG患者特异性生存期的危险因素,是卵巢癌患者预后不良的危险因素. CDCA5高表达是KIRP,KIPAN,LIHC,THCA,PAAD,PRAD和肉瘤样肺癌(SARC)患者无病生存期的危险因素,是UCS患者的保护因素.
-
研究检测了泛癌中CDCA5的表达与免疫细胞浸润之间的相关性. 使用TIMER 2.0数据库获得6种免疫细胞(B细胞、CD8+T细胞、CD4+T细胞、巨噬细胞、中性粒细胞和树突状细胞)的浸润评分,发现在多种恶性肿瘤中存在很强的相关性. KIRC,THCA,LIHC是相关性最高的3种,同时其线性回归图显示,CDCA5高表达与免疫细胞浸润量的增加相关. 值得注意的是,在LIHC中,CDCA5的表达与免疫细胞浸润量之间有更强的相关性(图 3). 树突状细胞在这3种恶性肿瘤中表现出最大的相关性. 使用3种不同的评分方式得到与CDCA5表达最显著相关的3种肿瘤分别为KIPAN,LUSC和THCA(免疫评分);KIPAN,LUSC和THCA(估计评分),BRCA,KIPAN和LUSC(基质评分)(图 4). 结果表明CDCA5的表达与KIPAN和LUSC的免疫浸润程度高度相关.
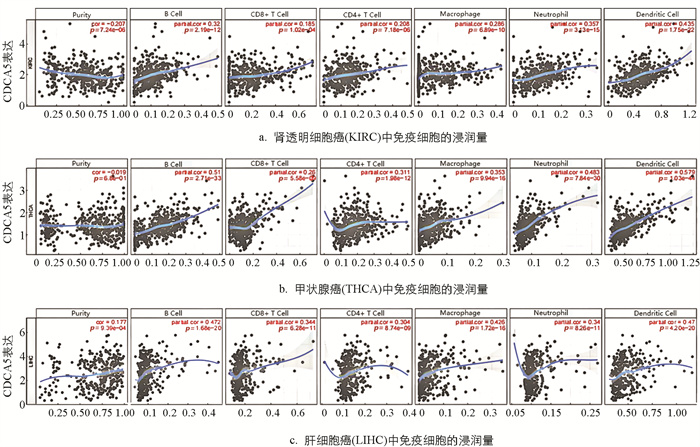
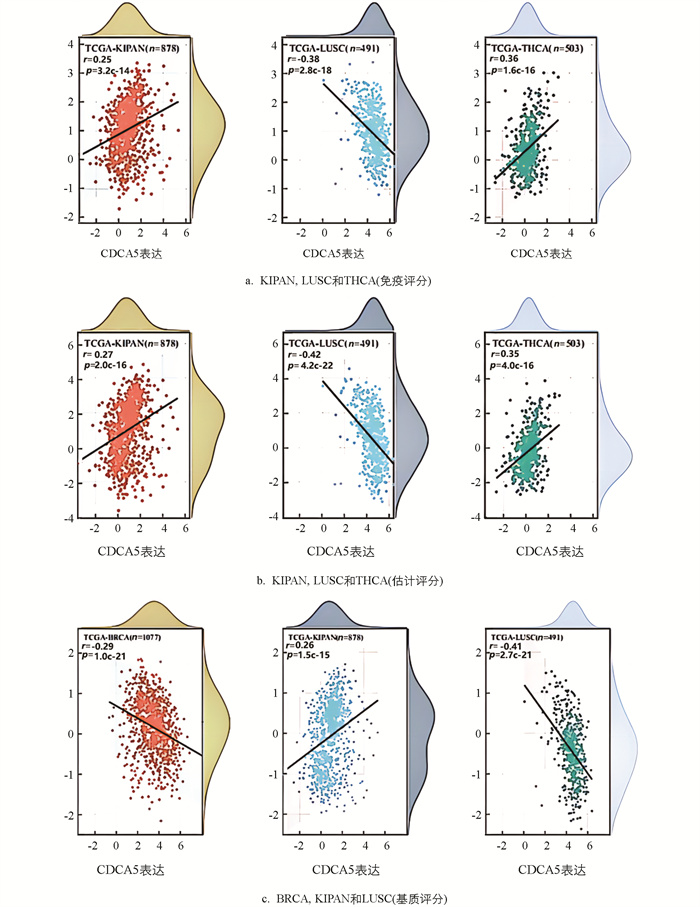
-
本研究分析了CDCA5与趋化因子基因、免疫刺激基因、免疫和受体基因之间的关系. 结果显示,CDCA5在泛癌中的表达与CCL17,CCL27和BTNL2基因的表达呈正相关. 此外,CDCA5的表达与MHC,HLA-G,CCRP,XCR1等受体相关.
-
基于GSCA网站,分析GDSC数据库结果显示,I-BEF-762,NPK76-Ⅱ-72-1,Navitoclax,Vorinostat和WZ3105对CDCA5表达的抑制率较高. CTRP数据库的分析结果显示,大多数化合物对CDCA5的抑制率较高,尤其是多西紫杉醇.
-
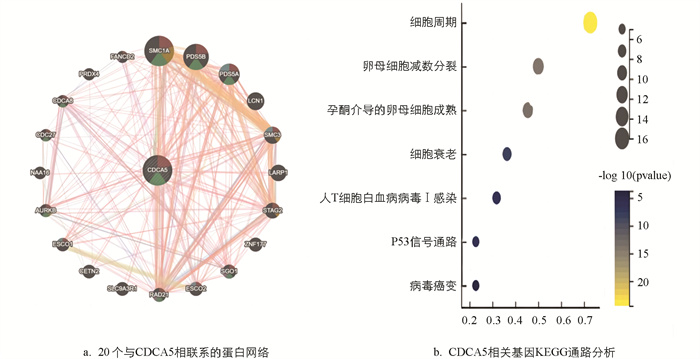
为了探究CDCA5在肿瘤发生癌变中发挥作用的可能过程,使用GeneMANIA在线程序为CDCA5创建了一个蛋白网络(图 5a). 20个来源于经实验验证的与CDCA5有密切联系的功能相似的基因位于外圆,而关键基因位于中心,显示CDCA5与SMC1A,PDS5B,RAD21,PDS5A和SMC3表现出显著的物理相互作用. 根据KEGG富集结果表明,“细胞周期” “卵母细胞减数分裂” “孕酮介导的卵母细胞成熟” “细胞衰老” “人T细胞白血病病毒Ⅰ感染” “P53信号通路”和“病毒癌变”可能参与了CDCA5在肿瘤致病中的作用(图 5b).
-
在泛癌中,CNV和CDCA5 mRNA表达之间存在Spearman关联. 在LUSC,BRCA,BLCA,OV,COAD,SKCM,LUAD和HNSC中,CNV与CDCA5 mRNA表达呈显著正相关;相反,这种相关性在PCPG,STAD,腱鞘巨细胞瘤(TGCT),UCEC,KIRC,ACC,KIRP,KICH,MESO,弥漫性大B细胞淋巴瘤(DLBC),急性髓系白血病(LAML),THCA,UVM,LGG,CHOL,THYM和LIHC中并不显著. 在大多数肿瘤中,甲基化与CDCA5 mRNA的表达呈负相关,这种相关性在TGCT,STAD,SKCM和BRCA中尤为明显.
-
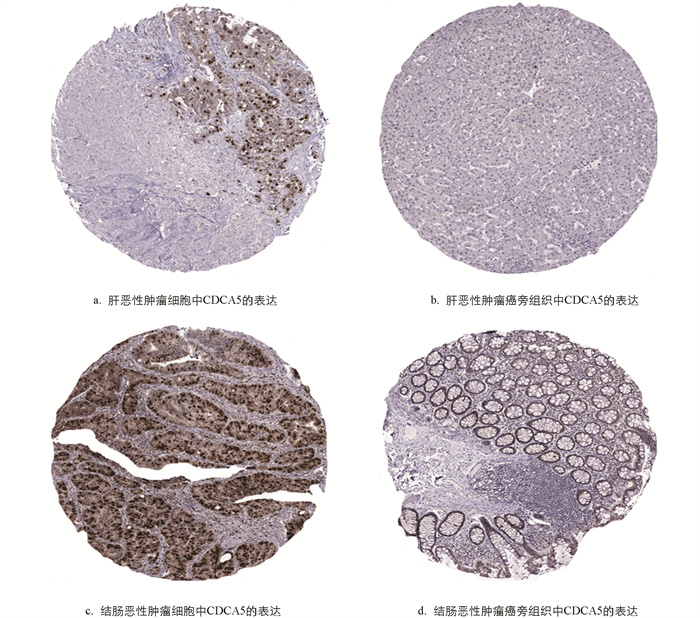
肿瘤样本与正常样本均来自于人类蛋白组图谱数据库,该数据库共包含13种不同人体组织的数据集. 使用免疫组化法验证CDCA5在肿瘤与癌旁组织中的表达,分析CDCA5在肝恶性肿瘤、结肠恶性肿瘤组织和癌旁组织中的表达情况. 在肝恶性肿瘤细胞中,绝大部分细胞呈中至强阳性胞质染色伴胞核染色,在癌旁组织中呈阴性;在结肠恶性肿瘤中,呈强阳性胞核染色,在癌旁组织中呈阴性(中倍放大). CDCA5在肝恶性肿瘤和结肠恶性肿瘤中的表达明显高于正常组织(图 6).
2.1. CDCA5在泛癌中的表达分析
2.2. CDCA5的表达与肿瘤分期呈正相关
2.3. CDCA5的预后意义
2.4. CDCA5表达与肿瘤免疫浸润水平的相关性研究
2.5. CDCA5表达与肿瘤免疫调控基因表达的相关性
2.6. CDCA5的表达与抗肿瘤药物的半抑制浓度(IC50)分析
2.7. 富集分析
2.8. CNV和甲基化与CDCA5 mRNA表达的相关性
2.9. 用免疫组化(IHC)检测CDCA5在肿瘤中的表达
-
姐妹染色单体的凝聚是细胞有丝分裂过程中染色体正确分离的关键,由黏合素复合体所介导[20]. CDCA5对于稳定黏合素复合物与DNA结合非常重要,以维持姐妹染色单体从S期到有丝分裂期的稳定黏合[21],因此,CDCA5对于遗传物质在子细胞中的正确分布至关重要. CDCA5和肿瘤之间的相关性已经证实,CDCA5在多种肿瘤中表达明显增高并且与肿瘤细胞增殖、转移和侵袭相关,其中包括膀胱癌[3]、肝癌[4-5]、胃癌[6]、肺癌[7]和乳腺癌[22-24]. 本研究的主要结果如下:1) 与正常组织相比,肿瘤中CDCA5的表达明显升高;2) CDCA5的高表达与肿瘤病理分期及预后不良相关;3) CDCA5的表达与肿瘤浸润水平相关;4) CDCA5的甲基化与肿瘤相关. 这些结果表明,CDCA5可作为肿瘤免疫浸润和预后不良的生物标志物.
富集分析显示,CDCA5的高表达主要与细胞分裂和细胞周期有关. 在DNA复制过程中,CDCA5被招募到染色质中,以确保姐妹染色单体的内聚,同时在有丝分裂期调控姐妹染色单体凝聚的消除[25]. CDCA5通过其C端的最后12个氨基酸与黏合素复合体的SA2亚基相互作用,进而稳定姐妹染色单体的内聚[26]. 不受控制的生长和侵袭被认为是最危险的病理改变[27]. DNA复制和黏合素复合体SMC3亚基的乙酰化共同促进了CDCA5与内聚复合体的结合,而WAPL可以逆转这一过程. CDCA5通过取代WAPL与PDS5结合来阻止WAPL,使DNA从黏合素复合体中分离出来,从而维持染色体的凝聚[28]. 在DNA断裂修复过程中,黏合素复合体被招募到DNA双链断裂位点,从而稳定姐妹染色单体凝聚. CDCA5在脊椎动物G2期稳定姐妹染色单体凝聚方面发挥着重要作用,当CDCA5从细胞中耗尽时,DNA双链断裂则无法修复,因此,CDCA5可能在DNA双链的修复中起辅助作用[1].
肿瘤浸润性免疫细胞是肿瘤微环境的重要组成部分,与肿瘤的发生发展及转移密切相关. 在正常情况下,免疫系统能够识别和中和肿瘤微环境中的肿瘤细胞,然而肿瘤细胞可能会利用多种生存生长和逃避机制来有效躲避免疫系统. 肿瘤免疫治疗可以恢复机体自然的抗肿瘤免疫反应,肿瘤浸润性免疫细胞对各种恶性肿瘤患者的预后有临床影响[29]. 研究表明,肿瘤浸润性免疫细胞会增加PD-1和PD-L1的表达,与肿瘤患者较差的预后和组织学分级相关[30]. CDCA5的表达与肿瘤纯度良好相关,但与免疫细胞浸润程度呈负相关,因此,CDCA5过表达与KIRC,THCA和LIHC的不良预后相关. 这些数据还表明,CDCA5的表达与肿瘤浸润水平有关. 在未来的研究中,我们可以通过检测CDCA5的表达水平来评价免疫治疗的效果,还可以开发针对CDCA5的靶向治疗,并可与传统的免疫治疗相结合,提高其疗效.
本研究还深入研究了CNV、甲基化与CDCA5 mRNA表达之间的关系. 此外,根据人类蛋白图谱,使用免疫组化方法验证了CDCA5在肝恶性肿瘤和结肠恶性肿瘤中的蛋白表达. 这些结果证实了肝恶性肿瘤和结肠恶性肿瘤的泛癌生物信息学分析结果的准确性和可靠性,我们将在更多的肿瘤中进行类似的分子生物学验证.
虽然CDCA5的表达与人类恶性肿瘤的免疫和临床生存相关,但尚不确定CDCA5是否一定通过免疫途径影响临床生存. 未来还需要进一步的体外或体内生物学实验加以验证,以增加治疗的有效性.
本研究对CDCA5进行了全面的评估,揭示了其作为患者预后指标的潜在作用及免疫调节作用. CDCA5是否可以作为肿瘤免疫治疗的靶点,有待进一步探索.
 DownLoad:
DownLoad: