


 下载:
下载:

-
大豆镰孢菌根腐病(Fusarium root rot)是一种分布范围广,危害重,防治困难的世界性土传真菌病害[1-2].研究发现,引起大豆根腐病的镰孢菌种类复杂,如尖孢镰孢菌Fusarium oxysporum、腐皮镰孢菌F. solani、禾谷镰孢菌F. graminearum、木贼镰孢菌F. equiseti、层生镰孢菌F. proliferatum、假禾谷镰孢菌F. pseudoproliferatum等,其中尖孢镰孢菌是主要致病菌之一,在各大豆栽培区普遍发生[3-5].课题组前期对四川不同生态区大豆根腐病致病镰孢菌进行分离鉴定发现,尖孢镰孢菌分离频率高且致病性强[6-7].
选育和利用抗病品种仍是防治大豆根腐病最有效的途径.我国虽然大豆资源丰富,但具有广谱、优质的抗(耐)病大豆品种仍较匮乏.近年来,东北地区围绕大豆镰孢菌根腐病病原菌种类及抗病种质资源鉴定开展了大量工作,获得了一批抗性表现良好的大豆品种及资源,并应用于生产及育种工作中[1, 8-10].国内外研究者还对250多份大豆材料进行了抗性鉴定,获得了抗性表现良好的镰孢菌主效数量性状位点(QTL)抗性位点[11].但关于镰孢菌侵染后大豆抗病反应的研究尚不足.
病原菌侵染会诱导植物体内一系列生理生化反应,如过氧化物酶活性(POD)、可溶性糖、丙二醛(MDA)、叶绿素和木质素含量等的动态变化.郑殿峰等[12]用尖孢镰孢菌侵染大豆根部后测定胚根中总糖含量、可溶性糖含量和MDA含量,认为可溶性糖含量和MDA含量可作为抗病育种和药剂调控与保护的研究靶标.张婵娟等[13]对大豆抗感品种进行菌液处理后测定其幼根内超氧化物歧化酶(SOD)、可溶性糖含量和MDA含量,抗感品种间的SOD、可溶性糖含量和MDA含量变化有明显差异.研究表明植物的抗病反应与氧化物酶活性POD、可溶性糖、MDA等有着重要的关系.研究受病原菌侵染后生理生化指标的变化可为大豆抗病品种选育和鉴定提供一定依据.
本试验以前期已分离鉴定的,引起四川大豆根腐病的致病尖孢镰孢菌Fusarium oxvsporum为病原菌材料,选用根腐病抗性不同的两个大豆品种,测定了尖孢镰孢菌接种后不同时间,大豆不同组织中过氧化物酶活性、可溶性糖、丙二醛、叶绿素和木质素含量等生理生化指标的变化情况,并研究其与大豆根腐病抗性的关系,为大豆根腐病抗病材料筛选与评价提供依据.
全文HTML
-
供试品种:南豆12和九月黄,均由四川农业大学农学院作物遗传育种实验室提供.
供试菌株:尖孢镰孢菌菌株Fusarium oxvsporum(Fo_Yaan06),由四川农业大学植物病理系前期分离鉴定,并保存在马铃薯葡萄糖琼脂培养基(PDA)上备用.
-
挑选健康大豆种子,依次用1% NaClO溶液处理2 min,75%酒精处理1 min,无菌水冲洗5~6次,放于装有灭菌蛭石的托盘中保湿催芽1 d,待用.病原菌接种参照Chang等[6]的带菌高粱粒接种法进行,放置于恒温培养箱中,(25±3) ℃,16 h光培养/8 h暗培养,于V1期即真叶完全展开,第一片复叶叶缘分离时,约15 d左右,进行发病情况调查.在接种后7 d和15 d收集大豆根组织为地下部分样品不同时间取样(7 d-SR和15 d-SR)、茎和叶部为地上部分(7 d-SL和15 d-SL),以生长7 d和15 d的未接种大豆地下部分(7 d-CSR和15 d-CSR)、地上部分茎和叶部(7 d-CSL和15 d-CSL)为对照.每个处理3次重复,每个重复9株大豆幼苗用于生理生化指标测定,试验重复3次.
-
大豆根腐病的病害分级标准参照王春玲等[8, 14]的方法,计算发病率(M)及病情指数(Id).大豆抗性类型参照杨帅[10]的方法分为5种:免疫(I),Id为0;高抗(HR),0<Id≤20;抗病(R),20<Id≤50;感病(S),50<Id≤80;高感(HS),Id>80.
-
于接种后不同时间,取大豆第一复叶叶片,剪成长、宽均为0.2~0.3 cm的叶段,测定各指标含量.叶绿素含量(mg/g)采用分光光度法测定[15-16];可溶性糖含量(μmol/mg)和丙二醛含量(μmol/mg)采用硫代巴比妥酸(Thiobarbituric acid,TBA)法测定[16];木质素含量(μmol/mg)采用紫外分光光度法测定[15-16].过氧化物酶(POD)活性测定:参照过氧化物酶(Peroxidase,POD)试剂盒(苏州科铭生物技术有限公司)说明书进行提取,并用紫外分光光度计测定A470下的吸光值,计算POD活性(U/g).
-
采用Microsoft Excel 2010进行数据整理与计算,采用SPSS 2.0软件进行差异统计学意义分析(统计学意义水平0.05).叶绿素含量、可溶性糖含量、MDA含量和木质素含量均为相对含量,即同一时间接种处理与未接种对照的含量比值(×100%). POD的相对活性以同一时间接种处理与未接种对照比值(×100%)计算.
1.1. 供试材料
1.2. 方法
1.2.1. 大豆催芽与接种方法
1.2.2. 病情调查及评级
1.2.3. 生理生化指标测定
1.2.4. 数据处理
-
接种尖孢镰孢菌15 d后大豆根腐病发生情况调查结果如表 1所示.南豆12和九月黄的发病率均为100%,但南豆12幼苗叶片正常,根部主根轻微变褐或变黑,但能通过侵染点继续生长,须根生长点变褐;而九月黄幼苗叶片发黄,较南豆12植株矮小,根部主根严重变黑,不能通过侵染点继续生长,须根明显减少,根部腐烂严重.依据杨帅[10]的抗性鉴定标准,南豆12为抗病品种,九月黄为感病品种.
-
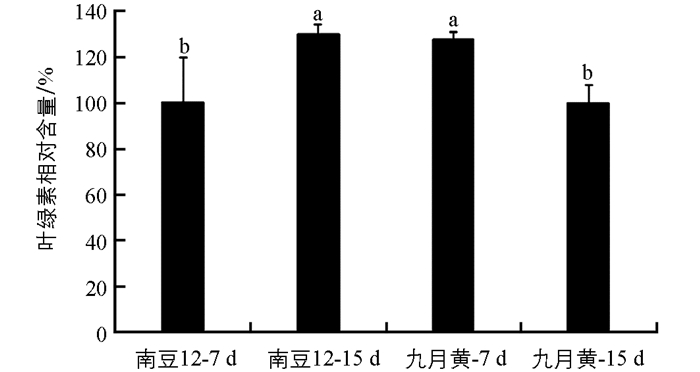
接种后测定两个大豆品种叶片的叶绿素含量,结果如图 1所示.接种7 d时,九月黄的叶绿素相对含量显著高于南豆12;而15 d时,南豆12叶绿素相对含量则显著高于九月黄.受尖孢镰孢菌侵染后南豆12叶片的叶绿素相对含量显著增加,而九月黄的叶绿素相对含量则显著减少.结果表明抗病品种受病原菌侵染影响小,可以正常生长;而感病品种的叶绿素减少,可能与病菌侵染后严重影响植株生长有关.
-
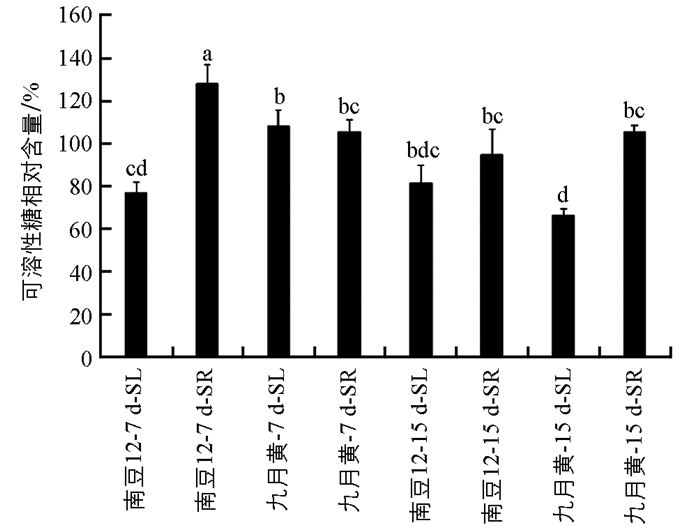
如图 2所示,接种尖孢镰孢菌后7 d和15 d时,大豆不同组织部位的可溶性糖相对含量变化不同.南豆12号地上部分(SL)的可溶性糖相对含量差异不具有统计学意义,地下部分(SR)的可溶性糖相对含量显著减少;而九月黄则地上部分可溶性糖显著减少,而地下部分差异不具有统计学意义.在接种7 d时,九月黄地上部分的可溶性糖相对含量显著多于南豆12,南豆12号地下部分显著多于九月黄;而在15 d时,两个品种的地上和地下部分差异均不具有统计学意义.结果表明在病原菌侵染前期,抗病品种的根部可溶性糖含量高,有利于增加抗性,而在后期病原菌积累量增加,导致抗病与感病品种无差异.
-
如图 3所示,接种后7 d和15 d时同一大豆品种不同组织中MDA相对含量变化不同.南豆12地上部分(SL)的MDA相对含量差异不具有统计学意义,而地下根部MDA相对含量显著降低;九月黄则地上部分和地下部分均明显降低.接种7 d时,两个品种地上部分MDA相对含量差异不具有统计学意义,而地下部分九月黄显著低于南豆12;在15 d时,南豆12地上部分MDA相对含量显著高于九月黄,而地下部分则差异不具有统计学意义.结果表明在病原菌侵染前期,抗病品种地下根部的MDA相对含量高于感病品种,表现出更强的抗性,而在侵染后期,伴随病原菌的大量增加,两个品种的MDA相对含量均受到影响.
-
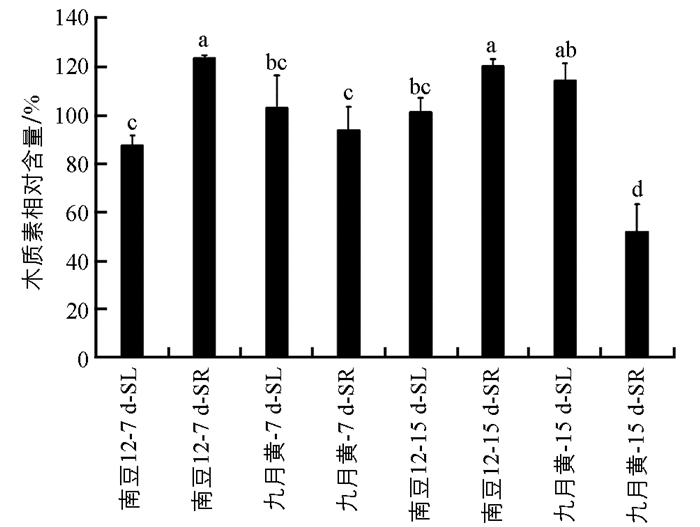
由图 4可见,在接种尖孢镰孢菌后7 d和15 d时,南豆12地上部分和地下部分的木质素相对含量变化差异均不具有统计学意义;九月黄地上部分差异不具有统计学意义,而地下部分差异具有统计学意义.接种后同一时间,两个品种地上部分的木质素相对含量差异均不具有统计学意义,而南豆12地下部分显著高于九月黄.结果表明抗病品种和感病品种根部木质素相对含量差异明显,抗病品种通过增加根部木质素含量以抵抗病原菌的侵入.
-
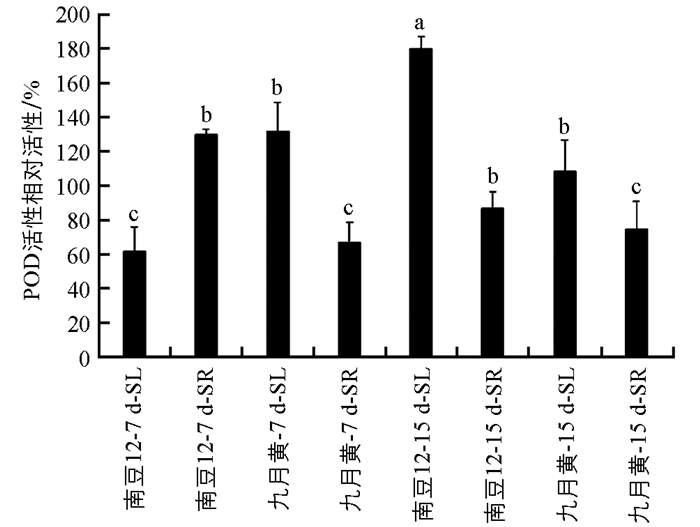
由图 5可见,在接种尖孢镰孢菌后的7 d和15 d时,南豆12地上部分POD相对活性均显著增加;地下部分POD相对活性差异不具有统计学意义;九月黄则地上部分和地下部分POD活性差异均不具有统计学意义.在接种7 d时,南豆12地上部分POD活性显著低于九月黄,地下部分POD活性显著高于九月黄;而在15 d时,南豆12地上部分和地下部分的POD活性均显著高于九月黄.结果表明在病原菌侵染过程中,品种的抗病性与POD活性有关.
2.1. 受尖孢镰孢菌侵染后两个大豆品种根腐病的发生情况
2.2. 受尖孢镰孢菌侵染后两个大豆品种叶绿素含量的变化情况
2.3. 受尖孢镰孢菌侵染后两个大豆品种可溶性糖含量的变化情况
2.4. 丙二醛(MDA)含量的变化情况
2.5. 木质素含量的变化情况
2.6. 过氧化物酶(POD)活性的变化情况
-
本试验以两个大豆根腐病抗性差异的品种南豆12和九月黄为材料,通过高粱粒接种测定了其地上茎叶组织和地下根部的叶绿素、可溶性糖、MDA、木质素含量和POD活性.结果发现:抗性品种的叶绿素相对含量受根腐病尖孢镰孢菌侵染后增加,而感病品种则降低,且抗性品种的叶绿素相对含量在侵染后期显著高于感病品种,这与欧志远[17]关于葡萄叶片内叶绿素的含量与霜霉病的抗性呈正相关相一致.受病原菌侵染后,抗感品种可溶性糖含量均减少,但感病品种减少幅度更大,表明病原菌侵染后大豆呼吸作用增强,与张婵娟等[13]的研究结果一致. MDA是膜脂过氧化的产物,病原菌侵入时需要突破细胞膜的屏障,而MDA的产生能对细胞膜造成伤害,加速病原菌入侵[18],导致抗性减弱.在本试验中,抗病品种和感病品种均以减少MDA含量来抑制病原菌的侵染,但在病原菌侵染前期,感病品种根部的MDA相对含量多于抗病品种,而在后期差异却不具有统计学意义,这与郑磊等[19]发现的在接种病原菌后,MDA含量的增长速度和积累量决定植株抗病能力的结论相似.木质素存在于植物的木质部中,可以增强植物体的机械强度,同时由于其疏水的化学特性,能使植物免受病原菌的侵害和扩展.本研究中随着接种时间的推移,抗病品种的木质素相对含量变化不明显但感病品种显著减少,这与魏丽娟[20]关于木质素在抗病和抗倒伏过程中起着重要作用的研究结果相似.同时,抗病品种地下根部的木质素含量显著高于感病品种,而茎和叶中的含量差异不大,推测可能与病原菌侵染及植株发病部位在根部,对地上部分影响较小有关. POD是一种关键的活性氧清除酶类,能保护膜结构,使植物具有一定的抗病性.本研究发现抗病性品种地上部分和地下部分的POD活性均明显高于感病性品种,与房玉林等[21]关于葡萄品种间的抗病性与POD活性关系的研究结果相似.
本试验以大豆根腐病的强致病菌尖孢镰孢菌为病原菌材料,研究大豆不同生理生化指标的变化,但田间引起大豆根腐病的镰孢菌种类复杂,后续将进一步鉴定不同大豆品种对多种根腐病致病菌的抗性,从而获得多抗大豆品种.不同大豆品种抗根腐病性能还与基因型有关,除生理生化指标外,将来可从分子生物学方面对抗病基因进行基因定位来探索其分子抗病机制[22],建立基于多个指标综合分析的大豆根腐病抗病性鉴定及评价方法[23].
本研究结果表明受根腐病致病尖孢镰孢菌侵染后,抗感病大豆品种的叶绿素含量、可溶性糖含量、丙二醛含量、木质素含量和POD活性变化存在差异,其中叶绿素、丙二醛、木质素、POD与品种抗性关系密切,可作为大豆根腐病品种抗性评价的重要指标及抗病育种的研究靶标.