


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
柑橘大实蝇(Bactrocera minax)是一种专门危害柑橘类果实的寡食性害虫,主要分布于中国、尼泊尔、印度及不丹等国,尤其在我国的为害最为严重[1-2]. 柑橘大实蝇是一化性昆虫,每年4~5月,成虫羽化出土,5~6月,雌成虫将卵产于果实内皮层,孵化为幼虫后就地取食危害果实;10月上旬开始,果实提前成熟并脱落,随后老熟幼虫钻出落果,钻入土壤中化蛹越冬[3-6]. 近年来,柑橘大实蝇对我国柑橘产业造成了巨大的经济损失,如2008年发生在广元的“蛆柑”事件,严重影响了全国的柑橘销售及社会大众的消费心理[7].
鉴于柑橘大实蝇对柑橘产业的巨大危害性,针对其综合防治的策略与方法已开展了大量研究[2, 5],目前主要防治手段包括:1) 果实套袋;2) 摘除虫果、捡拾落地果并集中处理;3) 深耕挖园灭越冬蛹;4) 移除虫源地柑橘树;5) 诱杀成虫;6) 释放不育成虫;7) 喷施化学药剂杀灭成虫. 农业防治与物理防治方法需要较高的人力和物力成本,不利于大规模推广;人工饲养该虫难度较大导致不育成虫的获取困难[3, 8];而诱杀成虫因具有简单、高效、对环境友好等特点逐渐受到重视,因此,越来越多针对柑橘大实蝇成虫的诱剂和设备得以开发利用,辅以传统的化学防治手段进行防控[9-13].
柑橘大实蝇成虫通过视觉定向,对黄绿色球形物体有强烈趋向性[14],根据这一特性而开发的球形诱捕器逐渐受到市场欢迎,且诱集效果已得到充分验证[11-12]. 然而,目前针对球形诱捕器防治效果的评价着重调查诱集到的成虫数量,这可能导致评价结果与实际防治效果存在偏差,因为所诱集到的成虫可能已经在果实内大量产卵,随后孵化为幼虫,因此,调查所诱集成虫的卵巢发育及产卵情况对全面评价球形诱捕器防治效果具有重要意义.
在本研究中,我们首次对球形诱捕器在不同时间及不同地点诱集的雌、雄成虫数量及雌成虫卵巢发育情况进行分析,以明确诱集效果及雌虫产卵情况,从而进一步判定球形诱捕器在田间的实际应用价值. 另外,研究结果还有助于明确柑橘大实蝇在田间的种群动态,为后续制定系统的柑橘大实蝇诱捕防治方案提供依据.
全文HTML
-
依科曼®新式球形诱捕器购自湖北谷瑞特生物技术有限公司,球体直径9±0.3 cm,表面附有青果绿色图层,均匀涂抹融合蛋白诱饵的高效粘虫胶,顶部有15~20 cm悬挂铁丝. 体视显微镜购自重庆奥特光学仪器有限责任公司.
-
于重庆开州选择一处柑橘园作为诱集试验果园. 根据果园的地形及环境,在果园内共设10个诱集点,其中果园边4个,果园内3个,近农舍果园边3个. 使用诱捕器时,将铁丝缠绕于柑橘树株高2/3处,尽量避免枝叶遮挡住球体,然后撕去包裹的油纸即可. 在2018年5月20日至7月7日期间,每3~4 d统计每个诱集球上的雌、雄成虫数量,收集的雌虫保存于70%的酒精中,并在同一位置更换新的诱集球. 酒精中保存的雌虫全部带回实验室,用于后续解剖观察.
-
雌虫带回实验室后,在体视显微镜下对每头雌虫进行解剖,观察卵巢发育并统计分级情况,卵巢分级参考刘颖等[15]制定的标准,略做修改,具体如下:
1) 卵巢发育初期(Ⅰ期):此时卵巢很小,呈乳白色絮状,卵巢长度约等于卵巢宽度. 无发育成熟的卵,尚未性成熟.
2) 卵黄沉积期(Ⅱ期):终端卵泡内卵黄开始累积,卵巢比Ⅰ级大,即卵巢长度比Ⅰ级长,卵巢宽度也比Ⅰ级宽. 无发育成熟的卵,尚未性成熟.
3) 成熟待产期(Ⅲ期):卵巢指数和卵泡长度与Ⅱ级相差不大,但所有形态特征比Ⅰ级、Ⅱ级更明显,其内卵黄含量充满整个终端卵泡体积的50%. 无发育成熟的卵,尚未性成熟.
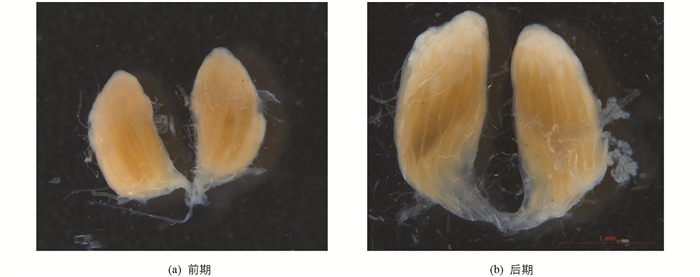
4) 产卵盛期(Ⅳ期):此等级所有指标都比其它等级大. 有发育成熟的卵,卵量最大,达到性成熟阶段. 此等级又分为前期和后期,前期未产卵或产卵量极少,卵巢形态饱满,无明显产卵痕迹;后期明显产卵,卵巢末端有产卵后形成的空壳,卵巢形态较为饱满(图 1).
5) 产卵末期(Ⅴ期):明显大量产卵,卵巢体积变小,形态不饱满,成熟卵的数量较少.
-
卵巢分级后,对性成熟期(Ⅳ期和Ⅴ期)卵巢中的成熟卵量进行统计,比较未明显产卵的卵巢(Ⅳ前期)与明显产卵的卵巢(Ⅳ后期-Ⅴ期)中成熟卵的数量.
-
采用SPSS 22.0软件进行单因素方差分析(one-way ANOVA)和LSD多重比较法对多组数据进行比较(p<0.05),两组数据采用独立样本t检验进行差异性分析(p<0.05),所有数据均以x ±s表示. 百分率数据在进行方差分析前进行反正弦平方根转换.
1.1. 材料
1.2. 诱集方法
1.3. 卵巢发育分级
1.4. 卵巢内成熟卵量统计
1.5. 数据分析
-
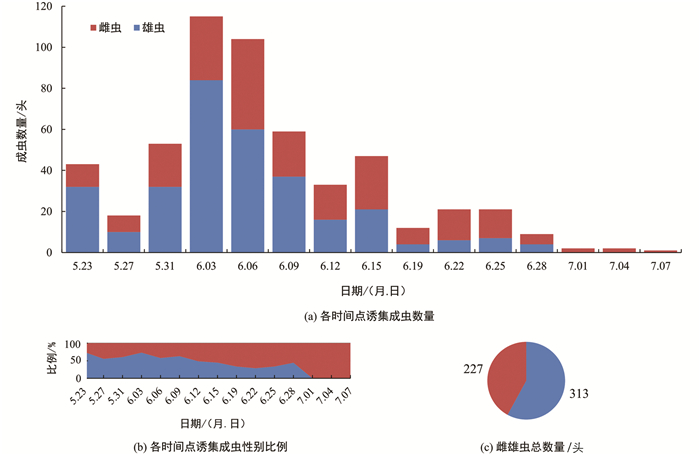
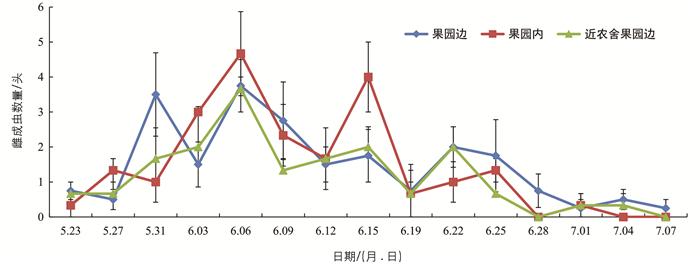
在5月20日至7月7日期间,一共收集样品15次,10个诱集点共收集成虫540头,其中雌虫227头,雄虫313头. 前期诱集雄虫较多,后期诱集雌虫较多,诱集的高峰期为6月初,6月中下旬以后诱集数量逐渐减少(图 2).
-
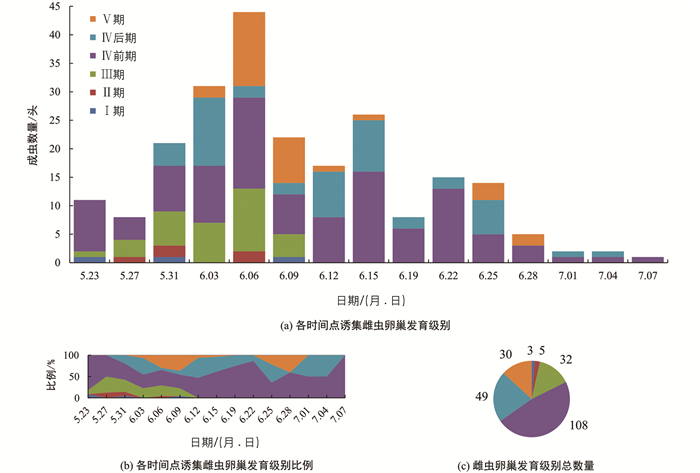
在诱集的所有雌虫中,148头(65.2%)未明显产卵(卵巢发育Ⅰ-Ⅳ前期),79头(34.8%)明显产卵(卵巢发育Ⅳ后期-Ⅴ期),其中30头(13.2%)大量产卵(卵巢发育Ⅴ期). 5月27日前诱集的所有雌虫均未明显产卵,6月9日前能诱集到未性成熟的雌虫,其后诱集的所有雌虫均已达到性成熟阶段(图 3).
-
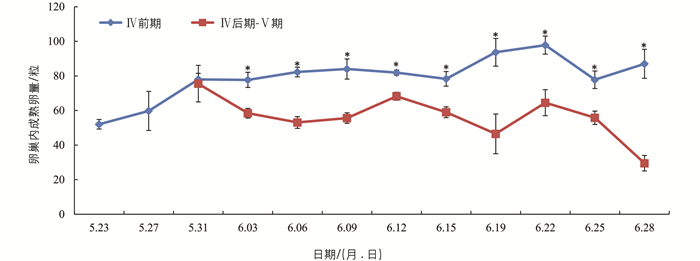
柑橘大实蝇卵巢发育至Ⅳ期达到性成熟,卵巢内有成熟卵. 随着时间的推移,未明显产卵的卵巢中,成熟卵数量逐渐上升,且都大于50粒/头;已明显产卵的卵巢中,成熟卵量逐渐下降,且都少于70粒/头. 6月3日以后,两者之间的差异有统计学意义(图 4).
-
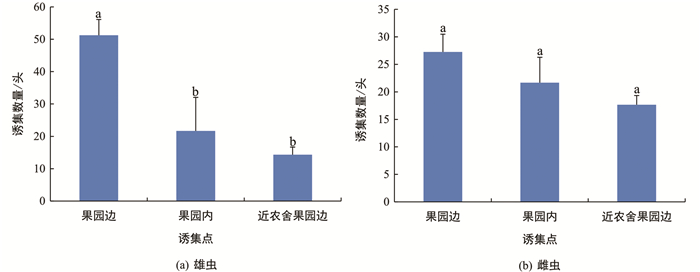
根据果园的地形及环境,共设置10个诱集点,其中果园边4个,果园内3个,近农舍果园边3个. 在5月20日至7月7日期间,果园边诱集点对雄虫的总诱集量为51.3头,高于其余诱集点,且差异有统计学意义;果园边诱集点对雌虫的总诱集量也高于其余诱集点,但差异无统计学意义(图 5).
果园边诱集点对雌成虫的诱集高峰期最早出现在5月31日;果园内和近农舍果园边诱集点对雌成虫的诱集高峰期最早出现在6月6日,此时果园边诱集点出现第2次诱集高峰,果园内诱集点在6月15日出现第2次诱集高峰(图 6).
-
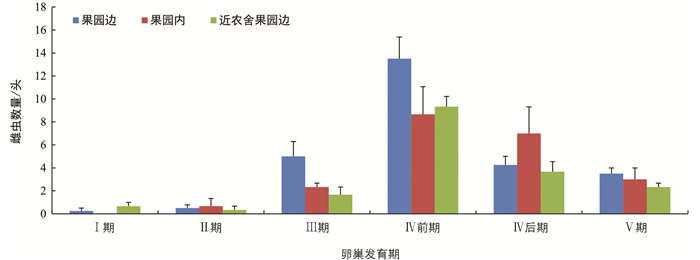
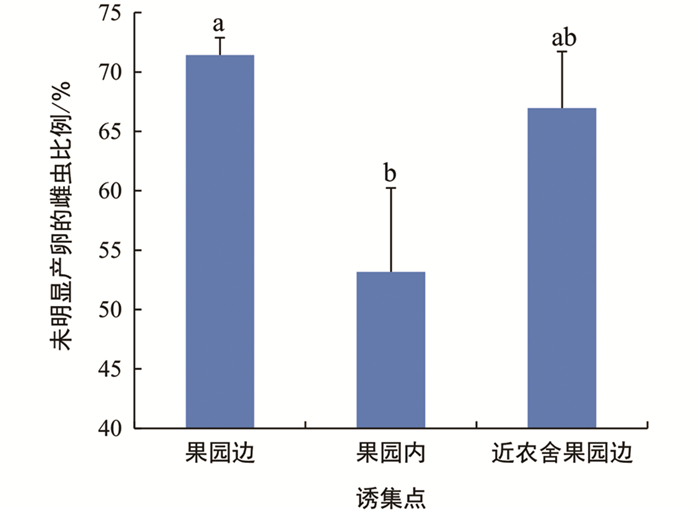
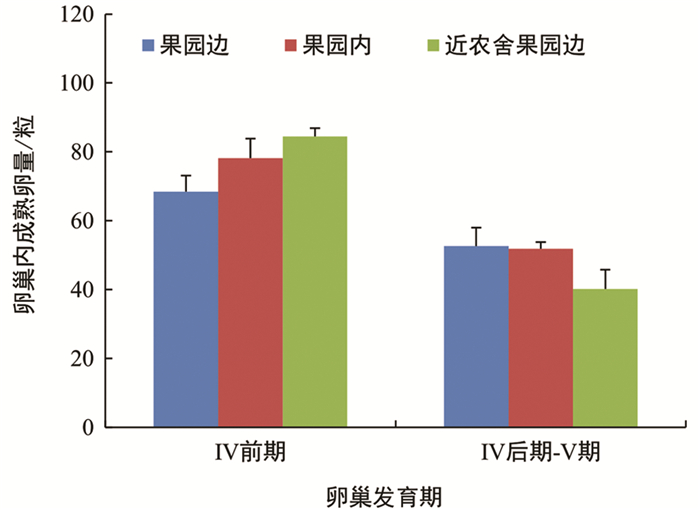
不同诱集点诱集的雌虫,卵巢发育有所差异,但差异无统计学意义(图 7). 然而,果园边诱集的雌虫中,未明显产卵的比例为71.43%,高于果园内诱集点(53.17%),差异有统计学意义;而与近农舍果园边诱集点(66.96%)相比,差异无统计学意义(图 8). 不同诱集点诱集到的雌虫卵巢中,成熟卵的数量差异无统计学意义(图 9).
2.1. 球形诱捕器在不同时间点诱集的柑橘大实蝇成虫数量
2.2. 球形诱捕器在不同时间点诱集的柑橘大实蝇雌成虫卵巢发育级别
2.3. 球形诱捕器在不同时间点诱集的柑橘大实蝇雌成虫卵巢内成熟卵数量
2.4. 球形诱捕器在不同诱集点诱集的成虫数量
2.5. 球形诱捕器在不同诱集点诱集的柑橘大实蝇雌成虫卵巢发育级别
-
在自然界中,实蝇成虫羽化后需要摄入花蜜、昆虫蜜露、花叶分泌物等多种食物为生殖发育提供营养物质[16]. 然而,不同的实蝇种类获取食物的途径存在较大差异,如昆士兰实蝇和桔小实蝇主要在寄主植物上获取食物[17-18],而柑橘大实蝇则在羽化后飞离果园,主要在非寄主植物上获取食物,待性成熟后再返回果园交配产卵[19],中国板栗(Castanea mollissima)花序对柑橘大实蝇成虫具有强烈的吸引力[20-21]. 本研究结果表明,依科曼®新式球形诱捕器对柑橘大实蝇雌、雄成虫都具有较好的吸引力,且前期诱集雄虫较多,后期诱集雌虫较多,7月以后只能诱集到雌虫,说明雄虫返回果园的时间早于雌虫,这可能与雄虫需要在交配前提早建立“领地”有关[19].
通过解剖诱集的雌虫,观察卵巢发育情况,发现大部分(65.2%)雌虫未产卵或极少量产卵,因此,在田间使用球形诱捕器可有效减少雌虫数量及产卵量,具有防治柑橘大实蝇的实际意义;而开始实施防治措施的最佳时期为5月底至6月初,此期间成虫羽化量逐渐增加,所有雌虫都未产卵或极少量产卵,其中部分雌成虫尚未性成熟,无产卵能力. 6月3日以后,诱集的雌虫已经性成熟,部分雌虫已开始产卵,且随着时间的推移,产卵量逐渐增大,在此阶段可结合其他措施进行防控,如食饵诱杀[13, 22]等. 此外,还可探索其他诱集手段,如性诱杀[23]、化学诱剂诱杀[20]等.
本研究所设的3个类型诱集点中,果园边诱集的成虫总数量、未明显产卵雌虫的数量和比例均高于果园内和近农舍果园边,且果园边最先出现诱集高峰,这与李杖黎等[24]的结果相似,表明了在果园边设置诱集点的重要性. 果园边诱捕成虫数量最多的原因可能是成虫从周边环境进入果园的过程中,首先会被果园边的诱捕器吸引. 果园边诱集的未明显产卵雌虫比例较高,可能是因为此时大部分雌虫尚未进入果园交配产卵. 当部分雌虫避开果园边诱捕器进入果园内部后,由于适合交配和产卵的地点显著增多,从而可能稀释了诱捕器的吸引作用,导致果园内的诱集高峰期延迟,且诱集的未明显产卵的雌虫比例下降. 近农舍果园边诱集点虽然也在果园边界,但是在观察诱集效果时,发现诱捕器上粘有大量与人类活动较密切的蛾类、蝇类等非目标昆虫,占据了大量的诱捕器表面,这表明人类活动可能对球形诱捕器的诱集效果存在干扰,导致诱捕的柑橘大实蝇数量减少.
综上所述,球形诱捕器对柑橘大实蝇雌、雄虫都有较好的诱集能力,且大部分诱集的雌虫未产卵或极少量产卵,具有用于田间防治的实际意义. 应用球形诱捕器防控柑橘大实蝇,应从成虫羽化初期开始,可适当增加果园边界的诱捕器密度,为成虫进入果园设置屏障,从而有效降低果园内的虫口密度. 此外,可结合处理虫果、食饵诱杀等其他防治手段,达到有效防控柑橘大实蝇、减轻危害程度的目的.