


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
自然界中生境复杂多变,生长在其中的植物通过多种性状的组合来适应其变化,植物对生境的适应也是一个复杂、综合的过程[1]. 由于不同生境内环境因子(光、热、水分、土壤等)存在差异,故导致不同生境内局部特殊的小气候形成,因此影响了植株对环境的适应程度[2]. 光照强度、水分、温度、CO2摩尔分数和土壤养分等环境因子均是影响光合作用的重要因素[3],而光合作用是影响植物生长发育的重要生理过程,光合生理特性在一定程度上能够体现植物对光照、CO2的利用特点及植物对生境的响应,表征植物对环境的适应性[4]. 研究表明,叶绿素质量分数、叶绿素荧光参数、光响应以及CO2响应参数均是评价植物生长的主要生理指标,对揭示植物在不同生境下的适应性具有重要作用[3, 5].
鸡爪槭(Acer palmatum Thunb.)是槭树科槭树属植物,树姿优美、叶形奇特,以其鲜艳丰富的叶色、稳定持久的观赏期,成为我国重要的园林观赏树种,深受人们喜爱,同时也弥补了传统绿化色彩单一、形式单调的缺点[6-7]. 我国虽然是槭树种质资源大国,但在观赏树木开发利用方面,与日本和欧美各国相比差距还很大[8-9]. 因此,加强对槭属优良品种资源的引种培育及生态适应性研究具有重要的经济价值和生态意义[6]. 目前,对槭属植物的研究主要集中在槭属叶色变化的种质资源、园林应用以及引种栽培等方面[6, 8],而关于不同生境之间的差异是否会对引种鸡爪槭的光合特性及叶色观赏质量产生影响,以及它们是如何响应环境的变化尚不清楚. 因此,本研究以4个典型生境内的不同品种鸡爪槭为试材,通过对不同生境下引种鸡爪槭光合特性和叶色的测定,比较不同生境下各品种的响应异同,以期从生理生态学角度揭示引起这种差异的原因,为鸡爪槭的栽培管理、引种驯化、不同环境下的响应方式和适生区选择提供理论参考.
全文HTML
-
结合前期对鸡爪槭实地考察及相关资料查阅,本试验选址于重庆市北碚区澄江镇造林地,该地属于典型的亚热带湿润季风性气候,气候温和,降雨充沛,全年日照总时数平均为1 285 h,最热月(8月)平均气温为28.2 ℃,极端最高温40 ℃;最冷月(1月)平均气温为3.1 ℃,极端最低温-4.6 ℃;年均降雨1 611 mm,无霜期317 d. 根据引种鸡爪槭在该区域种植分布情况,选取了4个具有代表性的生境,依次为低海拔阳坡(生境1,简称H1)、中海拔阳坡(生境2,简称H2)、高海拔阳坡(生境3,简称H3)及低海拔阴坡(生境4,简称H4),生境详细情况如表 1所示.
-
试验材料为引种于江西省宜黄县三农公司的3个品种鸡爪槭,分别为红舞姬HWJ、橙之梦CZM、三季黄枫SJHF. 3个品种嫁接苗(采用芽接法,砧木为1年生地径0.8mm左右的青枫)最初引种到本试验区的平均株高均为60.25±2.01 cm,平均地径均为12.45±1.16 mm,按照3 m×4 m的株行距进行定植,且采用统一的施肥、防虫和除草等管理措施. 试验开始于2021年3月上旬,依据随机性和独立性的样地选取原则,在每个生境内设置3个20 m×20 m的样方,每个样方之间至少相隔30 m,共计12个样方. 在每个样方内,不同品种鸡爪槭分别选取5株长势一致的样树进行挂牌标记,同时对4个生境中土壤养分、鸡爪槭叶色和光合指标进行测定.
-
每个样方按照梅花形布点方式选取5个采样点,用直径10 cm的土钻取0~20 cm表层土,混合均匀后带回试验室. 将采回的土样除去石块和动植物残体等杂物,经风干过筛后,根据《土壤农化分析与环境监测》中的方法测定土壤理化性质[10]. 其中,土壤pH值采用酸度计法测定;有机质(SOM)采用重铬酸钾容量法测定;全氮(TN)采用半微量凯氏法测定;碱解氮(AN)采用碱解扩散法测定;全磷(TP)采用氢氧化钠熔融-钼锑抗比色法测定;速效磷(AP)采用氟化铵盐酸浸提钼锑比色法测定;全钾(TK)采用氢氟酸高氯酸消煮火焰光度计测定;速效钾(AK)采用乙酸铵浸提火焰光度计法测定.
-
本试验使用的DEM数据为30 m的ASTER GDEM数据,该数据来源于中国科学院计算机网络信息中心国际科学数据镜像网站[11]. 以30 m的DEM为数据源,用SAGA GIS软件分别计算出每个生境的相对坡位指数(Relative Slope Position,RSP)、海拔(Elevation,EL)、坡向、坡度、沟谷深度(Valley Depth,VD)、地形湿度指数(Topographic Wetness Index,TWI)等地形因子.
-
采用Li-6800便携式光合测定仪调至叶绿素荧光模式,选取样树上最新完全展开的健康功能叶于2021年7月中旬9:00-11:30进行测定. 测定前用铝箔纸包裹叶片充分暗适应30 min,测得初始荧光(F0)、最大荧光(Fm)、最大可变荧光(Fv)、最大光化学效率(Fv/Fm),之后将待测叶片在目标光强下充分适应,测得稳态荧光(Fs)、电子传递效率(ETR)、实际光能捕获效率Y(Ⅱ),最后对叶片进行强饱和脉冲光激发,测得光化学猝灭系数(qP)和非光化学猝灭系数(NPQ).
叶绿素荧光测定完成后,立刻采摘叶片,用-80 ℃液氮低温保存,带回试验室参照《植物生理学实验教程》中的方法[12]测定光合色素:叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)、叶绿素(Chls).
于2021年3月10日(变色初期,Ⅰ)、3月25日(变色中期,Ⅱ)、4月10日(变色末期,Ⅲ)、7月25日(生长旺盛期,Ⅳ)、11月25日(生长末期,Ⅴ)采集叶片,对鸡爪槭叶色进行动态测定. 将清洗干净的每片叶子置于白色A4纸上,用EOS7D Mark Ⅱ数码相机进行拍照,在Adobe Photoshop CS6软件中进行分割,去除叶柄、托叶和白色背景,将保留的叶片以PNG文件格式存储,用Matlab 7.12.0软件编写代码提取颜色指数[13]:色彩亮度L*、红绿色度a*、黄蓝色度b*.
-
采用Li-6800便携式光合测定仪测定光响应曲线,于7月中旬9∶00-11∶30连续晴朗天气选择样树第4~5片功能叶进行测定. 设定参比室CO2摩尔分数为400 μmol/mol,温度为25 ℃,RH为65 %,气体流速为500 μmol/s,光合有效辐射(PAR)强度梯度为:1 800,1 500,1 000,600,300,200,100,80,50,20,0 μmol/(m2·s),测定前在1 200 μmol/(m2·s)的光强下诱导15 min,每2 min自动记录一个点. 每个样方重复3株,结果取平均值. 利用直角双曲线修正模型[14]进行拟合并计算出最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(Rd)、表观量子效率(AQY)和拟合决定系数(R2).
-
采用Li-6800便携式光合测定仪红蓝光源,于7月中旬选择连续晴朗天气,在9∶00-11∶30选取与测定光响应相同部位的叶片. 设置光强为1 800 μmol/(m2·s),温度为25 ℃,RH为65 %,气体流速为500 μmol/s,设定CO2摩尔分数梯度为:400,300,200,100,50,400,600,800,1 000,1 200,1 500,1 800 μmol/mol,每个样方重复3株,结果取平均值. 用直角双曲线修正模型[14]进行拟合并计算出初始羧化速率(α)、饱和胞间CO2摩尔分数(CSP)、CO2补偿点(CCP)、光呼吸速率(Rp).
-
使用Microsoft Excel 2010软件进行数据整理,采用SPSS 23.0单因素(one-way ANOVA)方差法分析了不同生境间环境因子以及不同生境下3种鸡爪槭叶色参数和光合特性的差异,采用双因素方差法分析了生境、品种及其交互作用对叶色参数、叶绿素荧光参数、光合特性参数的影响;环境因子与植物光合参数之间的关系采用冗余分析法分析;使用Origin 2018软件进行图表制作.
1.1. 研究区概况
1.2. 试验设计
1.3. 试验指标测定方法
1.3.1. 土壤样品的采集与相关指标的测定
1.3.2. 地形数据测定方法
1.3.3. 光合色素、叶绿素荧光及叶色参数的测定
1.3.4. 光响应曲线及光合参数测定
1.3.5. CO2响应曲线测定
1.4. 数据处理与统计
-
不同生境中土壤养分质量分数、地形因子、温度、相对湿度等均是影响植物光合生长的主要因子. 由表 1可知,生境中年平均温度H1最高、H3最低,坡度H4最高、H2最低,相对坡位指数从高到底依次为:H4,H2,H1,H3,土壤pH值从高到底依次为:H4,H1,H3,H2,且不同生境间差异有统计学意义;土壤中AN,AP从高到底依次为:H3,H2,H1,H4. H3,H2与H1,H4差异有统计学意义,但H1和H4之间差异无统计学意义;土壤含水率在不同生境之间从高到底依次为:H3,H2,H1,H4,且差异有统计学意义(p<0.05);不同生境间AK和TN差异无统计学意义(p>0.05).
-
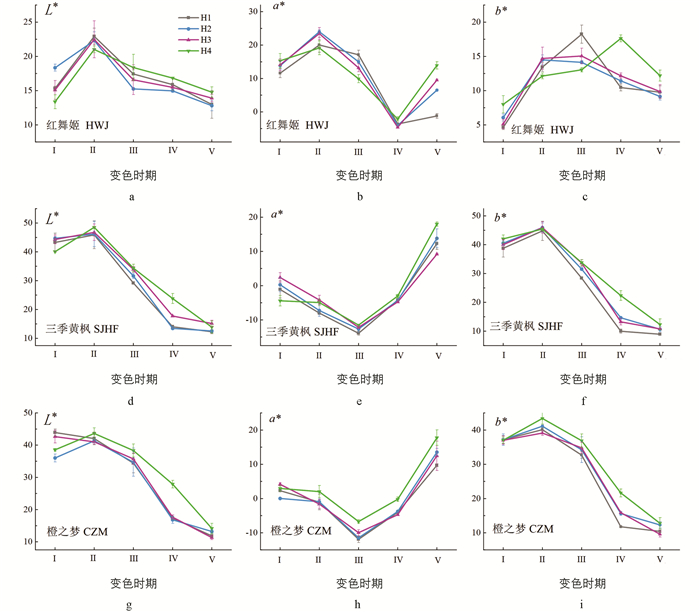
各处理叶色参数L*,a*,b*值随时间的变化趋势如图 1所示,在各生境中,3种鸡爪槭的L*值在变色中期最高,之后逐渐下降,其中变色中期至生长旺盛期下降趋势最大,生长末期达到最低值;三季黄枫、橙之梦的a*值在变色末期最低,之后逐渐上升,在生长旺盛期至生长末期上升趋势最大,b*值在变色中期达到最大值,之后逐渐下降,其中变色末期至生长旺盛期下降趋势最大;红舞姬的a*值在变色中期达到最大值,随后逐渐下降,在生长旺盛期达到最小,之后又趋于上升. 在变色中期,H2,H3中红舞姬的a*值显著高于其他生境a*值的平均值,其增量分别为2.35,1.77,相似海拔不同坡向(H1,H4)红舞姬a*,b*差异无统计学意义;三季黄枫在同一坡向不同海拔的b*值差异无统计学意义,分别为:44.77,45.90,45.92;橙之梦在相似海拔阴坡的b*值显著高于阳坡,且比平均值高2.46.
-
通过对不同生境下引种鸡爪槭光合色素进行分析发现(表 2),4个生境中,红舞姬、橙之梦Chla/Chlb差异无统计学意义(p>0.05),三季黄枫阴坡Chla/Chlb显著低于阳坡;红舞姬、三季黄枫的Chla,Chlb,Car和Chls质量分数从高到底依次为:H1,H3,H2,H4,其中H2与H3差异无统计学意义(p>0.05);橙之梦Chla,Chlb,Car和Chls质量分数从高到底依次为:H3,H2,H1,H4,且阴坡的Chla,Chlb,Car和Chls显著低于阳坡. 在H1中,橙之梦的Chla,Car和Chls显著低于红舞姬和三季黄枫(p<0.05),而3个品种的Chlb,Chla/Chlb差异无统计学意义;H2中橙之梦的Chla,Chlb,Car和Chls显著高于红舞姬和三季黄枫(p<0.05).
-
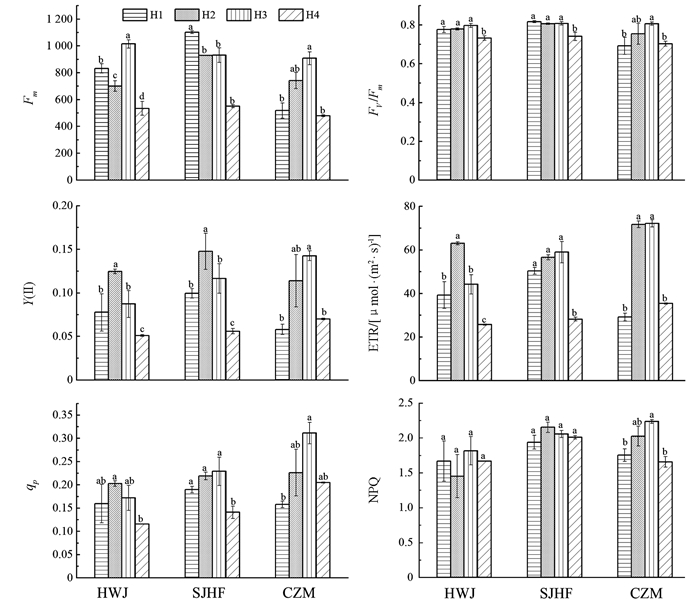
不同品种鸡爪槭的叶绿素荧光参数随着环境因子的不同而不同(图 2). 在不同生境中,红舞姬Fm从高到底依次为:H3,H1,H2,H4,且不同生境间差异有统计学意义(p<0.05);H4中Fv/Fm显著低于其他生境(p>0.05);Y(Ⅱ)、ETR和qp从高到底依次为:H2,H3,H1,H4,且H2显著高于其他生境(p<0.05),H3次之,H4最低. 三季黄枫在H1中Fm显著高于其他生境;阴坡中Fv/Fm、ETR、qp显著低于其他阳坡(p<0.05);H2中的Y(Ⅱ)显著高于其他生境;NPQ在H2中最大,H4中最小,分别为2.15,1.94;同一坡向不同海拔间Fv/Fm,ETR,qp,NPQ差异无统计学意义. 在不同生境中,橙之梦Fm,Fv/Fm,Y(Ⅱ)、NPQ、qp在H3中显著较高,分别为908.25,0.81,0.14,72.23,0.31;相似海拔不同坡向橙之梦Fm,Fv/Fm,Y(Ⅱ),ETR,NPQ差异无统计学意义. 由于不同品种鸡爪槭存在中间差异以及不同物种适应环境能力不同的现象,因此,不同生境下,3种鸡爪槭叶绿素荧光参数值随环境因子的改变升高或降低.
-
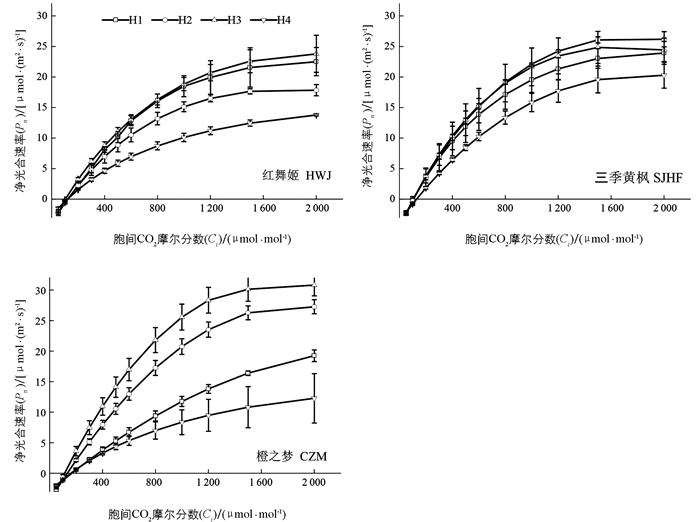
由图 3可知,引种鸡爪槭在不同生境内叶片Pn随PAR的变化趋势基本一致,当PAR值<300 μmol/(m2·s)时,3种鸡爪槭叶片Pn近似呈直线上升;当300<PAR值<600 μmol/(m2·s)时,Pn均随PAR上升而缓慢上升;当PAR值>600 μmol/(m2·s)后,Pn逐渐趋于平稳;当PAR值>1 500 μmol/(m2·s)后,三季黄枫Pn在H1,H2,H3略有上升,三季黄枫在4个生境中的Pn随PAR值的变化差异较小.
如表 3所示,在4个生境中,红舞姬Pnmax从高到底依次为H3,H1,H2,H4,且不同生境间差异有统计学意义;H1中LCP显著高于其他3个生境(p<0.05);Rd在H1中最大,H3次之,分别为:1.37,1.34 μmol/(m2·s). 同一坡向不同海拔三季黄枫LCP,AQY,Rd差异无统计学意义,相似海拔不同坡向三季黄枫LCP,AQY,Rd差异无统计学意义(p>0.05);Pnmax表现为H1最高,H3次之. 橙之梦Pnmax从大到小依次为:H3,H2,H1,H4,且阴坡显著低于阳坡(p<0.05);H1中LCP显著高于其他生境;AQY表现为H2,H3显著高于H1,H4(p<0.05),且相似海拔不同坡向差异无统计学意义. 在H1中,3种鸡爪槭AQY差异无统计学意义;三季黄枫Pnmax显著高于红舞姬、橙之梦;Rd从高到底依次为:橙之梦、三季黄枫、红舞姬. 在H2中,3种鸡爪槭的LCP和AQY差异无统计学意义,Pnmax从高到底依次为:三季黄枫、橙之梦、红舞姬,且差异有统计学意义(p<0.05). H3中各引种鸡爪槭Pnmax,LCP,AQY和Rd差异均无统计学意义(p>0.05).
-
由图 4可知,在不同生境中,当Ci<1 600 μmol/mol时,3种鸡爪槭Pn随Ci呈直线上升,且橙之梦在不同生境间Pn随Ci变化差异较大;当Ci>1 600 μmol/mol时,3种鸡爪槭Pn上升趋于缓慢,三季黄枫的Pn在4个生境间变化差异较小,而橙之梦3种鸡爪槭Pn在4个生境间变化较大.
由表 4可知,在4个生境中,红舞姬在H3中Anmax显著高于其他生境,而相似海拔不同坡向Anmax差异无统计学意义;阴坡α和Rp显著低于阳坡,而同一坡向不同海拔间差异无统计学意义. CSP差异无统计学意义(p>0.05). 三季黄枫在4个生境中的CCP,CSP和Rp差异无统计学意义;阴坡Anmax显著低于阳坡(p<0.05). 橙之梦的α,Anmax在H2和H3中较高,但差异无统计学意义;α,CSP和Rp在相似海拔不同坡向间差异无统计学意义(p<0.05). 在H1中,3种鸡爪槭的α,Anmax差异无统计学意义,橙之梦CSP和CCP显著高于其他2种鸡爪槭,而Rp显著低于其他2种鸡爪槭(p<0.05). 在H2中,3种鸡爪槭的α,CSP和Rp差异无统计学意义(p>0.05). 在H3中,3种鸡爪槭的Rp差异无统计学意义(p>0.05),橙之梦的Anmax显著高于红舞姬、三季黄枫(p<0.05). 在H4中,3种鸡爪槭的α,Anmax,CSP差异无统计学意义(p>0.05).
-
表 5为生境、品种及其交互作用对引种鸡爪槭叶色、光合参数的双因素方差分析结果. 由表 5可知,品种的不同对叶色参数L*,a*,b*值产生极显著影响,同时,生境与品种的交互作用对a*,b*值产生显著影响. 生境变化对Chla,Chlb,Chls,Car,LSP,AQY,CSP,qp,qN产生极显著影响(p<0.01),对α,LCP和CCP无显著影响;各品种间Pnmax,Anmax,Fv/Fm,ETR差异有统计学意义(p<0.05);同时,生境与品种的交互作用对Chla,Chlb,Chls,Car,LSP,AQY,CSP,Pnmax,Anmax,Fv/Fm,ETR,qp,qN分别产生极显著影响(p<0.01),对LCP产生显著影响,而α,CCP不受生境变化及其与品种的交互影响(p>0.05). 总体来看,叶色参数与不同生境间无显著影响,而品种间叶色参数差异极有统计学意义;生境及其与品种的交互作用对绝大多数光合参数都造成了显著或极显著影响.
-
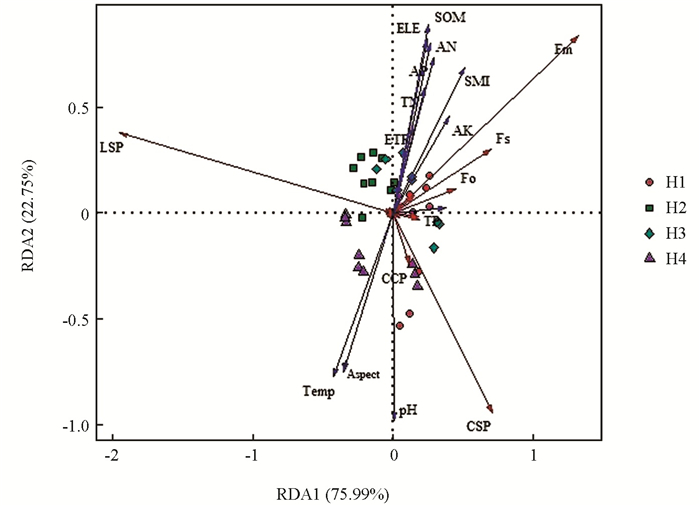
本研究采用冗余分析方法,描述了环境因子与引种鸡爪槭光合参数之间的相互关系,由表 6可知,第1轴和第2轴的特征值分别为0.759 9和0.227 5,前两轴环境因子对光合特性参数的累计解释量达到98.74 %,说明排序结果可信,能较好地解释两组变量之间的关系. 由图 5可知,环境因子的贡献率排在前7位的依次为:pH值、土壤含水率、海拔、坡向、有机质、碱解氮、平均温度,其中土壤pH值、坡向和温度与CSP呈显著正相关;有机质、碱解氮、土壤含水率、速效钾与F0,Fm,Fs,ETR呈显著正相关,与LSP,CSP呈显著负相关. 综上,环境因子与光合参数之间有很强的相关关系.
2.1. 4个生境环境因子分析
2.2. 不同生境对引种鸡爪槭叶色参数的影响
2.3. 不同生境对引种鸡爪槭光合特性的影响
2.3.1. 不同生境对引种鸡爪槭叶片光合色素质量分数的影响
2.3.2. 不同生境对引种鸡爪槭叶绿素荧光参数的影响
2.3.3. 不同生境对引种鸡爪槭光响应参数的影响
2.3.4. 不同生境对引种鸡爪槭CO2响应参数的影响
2.4. 生境、品种及其交互作用对引种鸡爪槭叶色、光合参数的影响
2.5. 引种鸡爪槭光合特性与环境因子的冗余分析
-
叶色变化的内部原因主要是光合产物的变化,引起植物体内各类色素及其比例发生变化,从而使叶片呈现出不同的色彩[15]. 而CIE L*a*b*是由国际照明委员会(Commission Internationale de L'Eclairage,CIE)于1931年建立的一种基于生理特性,与设备无关的色彩系统. 其中L*值为亮度参数,当颜色由黑变白时,对应的L*值由0变为100;色相a*表示颜色的红/绿程度,a*值越大,叶片越红;色相b*表示颜色的黄/蓝程度,b*值越大,叶片越黄[16]. 本研究中,在变色中期,不同生境下红舞姬a*值由3月变色中期的最大值到7月生长旺盛期的最小值,之后又趋于上升,叶片颜色变化为:亮红色—深绿色—暗红色;在变色中期,不同生境下橙之梦、三季黄枫的b*值达到最大值,在生长末期,a*值达到最大值,可见,在一个生长季中,橙之梦、三季黄枫叶片颜色变化为:浅黄绿色—深绿色—暗红色.
光照能显著影响植物叶片中的色素合成和相对质量分数,植物光合色素质量分数的差异又可以反映植物的生长状况和光合作用的强弱[17-18]. 研究表明,合成叶绿素的相关基因会通过质体信号表达影响植物光合作用,Chls质量分数越高,越有利于植物在低光强下捕捉光能,从而提高光合作用,积累更多的有机物[19-21],并且叶片中较高的Chls,Chla和Chlb与其高光合能力相对应[22],Chla和Chlb的质量分数可以反映叶片光能利用效率和光能捕获能力[23]. 本研究中,3种鸡爪槭在阴坡中Chls,Chla,Chlb显著低于阳坡,表明阴坡上生长的鸡爪槭适应光照强度变化的能力弱,从而光合能力差. 本研究发现,不同生境下各品种鸡爪槭AQY在0.05~0.10之间,与一般植物在适宜生长条件下的AQY(0.03~0.05)相比,略有偏高[24-25]. Fv/Fm反映PSII利用光能的能力,一般维持在0.75~0.85,该值下降是植物受光抑制最明显的特征[26-28]. 本研究中,H1,H4橙之梦,H4红舞姬的Fv/Fm均低于0.75,结合AQY值,说明在该生境下红舞姬、橙之梦PSII反应中心受到抑制,潜在活力降低,生长受到影响. Anmax是植物在CO2饱和点处达到的最大净光合速率[29-30]. Rp是一种消耗过剩光能、保护光合机构免于强光破坏的指标[31-32]. 研究发现[33],羧化速率与RuBPCase酶的活性呈正相关,Anmax和α较高的植物其叶片1,5-二磷酸核酮糖(RuBP)的再生能力强. 在同一生境下,三季黄枫的Pnmax,Anmax,α,Rp,AQY和Fv/Fm均高于红舞姬、橙之梦,说明三季黄枫对CO2的同化能力和利用效率高,其自身具有高光合潜能,可以进行更多光合物质生产与积累,可以认为三季黄枫在各生境下均可正常生长,且适应环境能力强于红舞姬、橙之梦,其原因可能是三季黄枫在适应环境过程中逐渐形成适于自身生长发育的生理生态特性.
叶绿素荧光参数是一组用于描述植物光合生理状况和光合作用机理的变量或常数值,反映了植物PSII的电子传递活性越大,光能利用率越大[34-35];NPQ是PSII反应中心以热形式耗散的过剩光能[36-38]. 本研究结果表明,生长于阴坡(H4)中的3种鸡爪槭与低海拔(H1)橙之梦的qP低于其他生境生长的苗木,说明阴坡中各品种鸡爪槭与低海拔中橙之梦PSII的电子传递活性低,即光合活性低,这与本试验AQY,Fv/Fm的结果一致;不同生境下红舞姬、三季黄枫的NPQ差异无统计学意义,橙之梦在H1,H4中的NPQ显著低于H2,H3,说明在不同生境下红舞姬、三季黄枫的热耗散或光保护能力相当,而橙之梦在H1,H4中的热耗散能力较弱.
环境因子间通常是相互影响的,并给植物生长和生理带来不同程度的效应[39]. 本研究中,Chla,Chlb,Chls,Car,LSP,AQY,CSP,Pnmax,Anmax,Fv/Fm,ETR受生境及生境与品种的交互影响极显著,说明上述生理指标对环境依赖性高于其他生理参数,且不同物种对环境的响应并不一致. 环境因子包括海拔、光照、土壤、气温、湿度等的变化影响植物与环境之间的水、气和养分的交换,进而影响植株的光合作用,最终对植物的生长发育产生影响[1, 40]. 坡向和坡度是重要的地形因子,其调控水、热组合,并且坡度能影响土壤水分渗透和地表径流,进而影响植株对环境水分的吸收[41-42]. RDA分析表明,环境因子中pH值、土壤含水率、海拔、坡向对鸡爪槭光合特征参数的解释量达到77.95%,说明它们是影响鸡爪槭光合特性的关键因子. 本研究发现,H4(阴坡)土壤pH值为8.20、坡度为32°,各引种鸡爪槭表现出较弱的光合作用,说明碱性土壤对引种鸡爪槭光合活性具有抑制作用,可能是土壤pH值影响了土壤养分的转化、释放、有效性及植物根系吸收养分的能力[43],同时,坡向与坡度共同作用影响了地表径流,进而影响了土壤含水率,对植物光合作用产生影响;而H3土壤pH值为弱酸性,海拔高、坡度缓,土壤中含有较高的氮素和磷素,这可能是3种鸡爪槭光合能力(Pnmax,ETR)显著提升的主要生理原因,使H3中各引种鸡爪槭均表现出较好的适应性.
-
生境差异对引种鸡爪槭的光合特性有显著影响,而环境因子中pH值、土壤含水率、海拔、坡向对鸡爪槭光合特性起主导作用. 阴坡中各鸡爪槭的光合能力均较弱,由此可知,引种鸡爪槭不宜生长于含水率低、坡度陡的碱性阴坡土壤中;而H2和H3的土壤为弱酸性,且坡度缓、土壤含水率高,3种鸡爪槭均表现出较好的适应性. 在同一生境下,三季黄枫的Pnmax,Anmax,α,Rp,AQY和Fv/Fm均高于红舞姬、橙之梦,表明三季黄枫捕获、传递光能以及进行能量转化的光合色素质量分数均大于其他2个品种;橙之梦在H1中AQY和Fv/Fm低于其他2个品种,说明橙之梦在中性、弱碱性的土壤中光合特性会受到影响,在酸性土壤中表现出较好的适应性,因此,可以认为三季黄枫生态幅宽且适应环境能力强,红舞姬次之,橙之梦最弱,适应性强的植物的光合速率高,光合产物积累量随之增加,有利于三季黄枫光合能力的提升,从而在竞争中保持优势地位. 不同生境下3种鸡爪槭均表现出较好的呈色.