


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
生长素响应因子(Auxin response factors,ARFs)属于植物B3超家族的一个亚家族,是一种植物特异性转录因子[1].ARF蛋白一般含有2个蛋白保守结构域,包括N端的B3结合域和C端的Aux/IAA结构域[2].目前,ARF基因家族在许多植物中已经被鉴定,包括拟南芥[3]、玉米[4]、水稻[5]、大麦[6]等,此外,小麦ARF基因家族也已经被众多学者进行过系统的研究与分析[7-9].这些工作对植物ARF基因的功能研究起到了重要的推动作用.
近年来,已经有大量的ARF基因在植物中被成功克隆.在模式植物拟南芥(Arabidopsis thaliana)中,AtARF5是第1个通过图谱克隆分离出的植物ARF基因,它被证实在植物的胚和维管组织的形成中发挥着重要作用[10].AtARF1和AtARF2的基因位点突变会影响雌蕊的生长发育,同时也会导致叶片衰老、花器官异常等现象的发生[11].AtARF3和AtARF4在植物生殖生长和营养生长中发挥着重要作用[12].AtARF7和AtARF19可以促进侧根形成,在激素信号通路中发挥重要作用[13].小麦TaARF20基因作为调控穗型、穗大小或穗粒数的关键基因,在小麦穗生长发育中发挥重要的调控作用[9].在水稻(Oryza sativa L.)中,OsARF1表达水平的降低会导致水稻株系活力低、生长迟缓且不育、叶片卷曲变短等,由此可见,OsARF1也参与了植物的营养生长和生殖生长[14].此外,ARF基因可以通过受到miRNA调控来发挥作用,研究发现,OsmiR167a可以通过调控生长素在茎部的不对称分布,抑制其靶基因OsARF12,OsARF17和OsARF25的表达,控制水稻分蘖角度[15].在转基因水稻中,过表达miR167会导致4个OsARF基因(OsARF6,OsARF12,OsARF17和OsARF25)的表达水平大幅减少,植株分蘖数显著减少[16].截至目前,关于ARF基因的功能研究主要还是以拟南芥为主,因此ARF基因家族的功能研究还有很多的工作要做.
小麦是我国最重要的粮食作物之一,基因组十分复杂.本研究在前人研究的基础上,对小麦TaARF14基因进行了克隆和表达模式分析,旨在为后续研究TaARF14基因功能提供重要的参考信息.
全文HTML
-
本研究使用的小麦材料为半冬性品种‘尚农6’(豫审麦20220077),于2022-2023年度种植于商丘市农林科学院双八镇试验基地(北纬34°53′,东经115°71′),小麦种植间距和肥水管理按照常规田间管理措施执行[17].2023年3月取处于小麦幼穗分化期(雌雄蕊分化期)的根、茎、叶、幼穗原基,4月取处于抽穗期的雄蕊、雌蕊等组织样品,取样后在液氮中速冻保存以备RNA提取,用于小麦转录因子TaARF14基因的克隆与分析.
-
采用Trizol法提取总RNA[18].所有RNA用1%的琼脂糖凝胶电泳进行质量检测,然后用Nanodrop ND-1000分光光度计测定浓度并进行定量,每个样本使用1 μg RNA进行反转录,cDNA第1链的合成使用南京诺唯赞生物科技有限公司的HiScript® Ⅱ 1st strand cDNA synthesis试剂盒进行,反转录得到的cDNA保存于-20 ℃冰箱中,用于小麦TaARF14基因的克隆和基因表达模式分析.
-
根据前人对小麦ARF基因家族的鉴定结果[7],TaARF14基因共包含3个典型的同源基因(转录本ID为TraesCS1A02G334900.1,TraesCS1B02G347900.2,TraesCS1D02G337400.1),根据其在染色体上的位置信息,分别命名为TaARF14-1A,TaARF14-1B和TaARF14-1D.从植物基因组数据库Ensemble Plants (
http://plants.ensembl.org/ )中分别获取TaARF14-1A,TaARF14-1B和TaARF14-1D的基因编码区序列和蛋白序列;使用DNAman 6.0(http://www.dnaman.com.cn/ )进行基因编码区序列的比对,通过SMART网站(http://smart.embl-heidelberg.de/ )鉴定蛋白序列的保守结构域;使用HMMER 3.3[19](http://www.hmmer.org/ )在植物基因组中进行TaARF14的同源基因比对,对筛选得到的氨基酸序列通过MEGA 7.0(http://www.megasoftware.net/ )进行多重序列比对;用邻接法(Neighbor-Joining method)构建系统发育进化树,参数选择默认. -
使用Primer premier 5.0(
http://www.premierbiosoft.com/primerdesign/index.html )分别设计PCR引物进行TaARF14-1A(正向引物序列:5′-GGGGCAAGCAAACAAAGA-3′;反向引物序列:5′-CCACGCAAGTGAATGTCAAGTA -3′),TaARF14-1B/1D(正向引物序列:5′-ATGGGGATCGACCTGAACAGC-3′;反向引物序列:5′-TCACATCCCCAGGGGAGCGGC-3′)基因编码区序列的扩增,将根、茎、叶、幼穗、雄蕊以及雌蕊的cDNA样本等量混合后作为模板进行PCR.采用上海翌圣生物科技股份有限公司的2×Hieff® PCR Master Mix(With Dye)配置PCR反应体系,PCR扩增程序为:95 ℃预变性5 min;95℃变性30 s,60 ℃复性1 min,72 ℃延伸1 min,35次循环;最后在72 ℃下延伸5 min,4 ℃保存.PCR产物经1%琼脂糖凝胶电泳后回收目的片段并进行测序,测序由北京六合华大基因科技有限公司完成. -
选择饱满、健康的‘尚农6’籽粒置于干净的培养皿中萌发,24 h后,将萌发的籽粒种植于营养土中并转移至培养箱进行培养,待小麦生长到3叶期时进行激素处理,分别喷施10 μmol/L生长素(IAA)溶液、200 μmol/L赤霉素(GA)溶液、100 μmol/L脱落酸(ABA)溶液,直至叶片全部湿润,分别在激素处理后0,1,2,4 h对幼苗的根、茎和叶片进行取样,取样后立刻在液氮中保存,随后在实验室提取所有样品的RNA,总RNA的提取和cDNA的合成参考1.2.
-
考虑到小麦中的3个同源基因具有相似的表达水平[20],我们根据TaARF14-1A,TaARF14-1B和TaARF14-1D的序列比对结果,在3个基因的同源片段设计通用引物进行TaARF14基因的表达分析.使用Primer premier 5.0(
http://www.premierbiosoft.com/primerdesign/index.html )在基因的保守序列设计TaARF14的qRT-PCR引物(正向引物序列:5′-GTCAACCTCGTAGGCATCTT-3′,反向引物序列:5′-CTCTCCGTACACCCAATCTT-3′),使用β-Actin(正向引物序列:5′-CCAAGGCGGAGTACGATGAGTCT-3′,反向引物序列:5′-TTCATACAGCAGGCAAGCACCAT-3′)作为内参基因.采用南京诺唯赞生物科技有限公司的ChamQ SYBR qPCR Master Mix配置qRT-PCR反应体系,反应体系参照qRT-PCR程序说明书执行,每个反应包含3次生物学重复,数据分析采用2-ΔΔCT法[21]进行计算和处理.
1.1. 试验材料
1.2. 总RNA的提取和cDNA的合成
1.3. 小麦TaARF14基因的生物信息学分析
1.4. 小麦TaARF14基因的克隆
1.5. 激素胁迫处理
1.6. 小麦TaARF14基因的表达模式分析
-
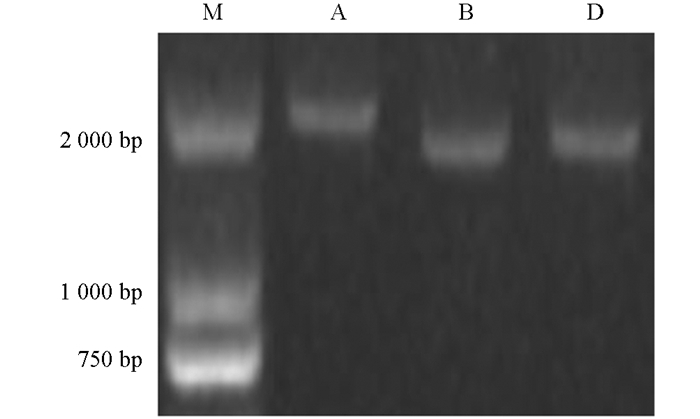
以‘尚农6’的各组织或器官的混合cDNA为模板,对TaARF14进行基因克隆,PCR扩增结果显示分别在2 551,2 097,2 097 bp处得到与目标片段长度相符的条带(图 1),其中TaARF14-1B,TaARF14-1D所用为相同的引物对.对扩增产物进行胶回收并测序后发现,TaARF14-1A,TaARF14-1B,TaARF14-1D与TraesCS1A02G334900.1,TraesCS1B02G347900.2和TraesCS1D02G337400.1的编码区序列完全相同.
-
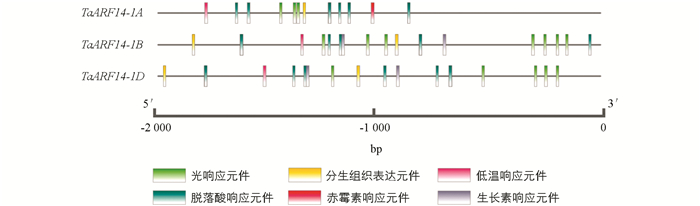
对编码序列进行序列比对后发现,3个同源基因(TaARF14-1A,TaARF14-1B和TaARF14-1D)的核酸序列具有极高的相似性,3个基因都具有10个外显子区域,且TaARF14-1A和TaARF14-1B基因的核酸序列相似性为95.13%,TaARF14-1A和TaARF14-1D基因的核酸序列相似性为94.94%,TaARF14-1B和TaARF14-1D基因的核酸序列相似性为96.07%.对蛋白序列进行比对发现(图 2),TaARF14-1A,TaARF14-1B和TaARF14-1D分别编码707,698和698个氨基酸,3条蛋白序列之间的差异氨基酸主要存在于C端,TaARF14的蛋白序列有B3和Auxin-resp两个保守结构域,其中B3结构域的氨基酸序列并未发生任何改变,Auxin-resp结构域共有4个差异氨基酸,结构域上部分氨基酸编码的差异可能会导致3个基因在基因功能以及基因表达水平上具有一定的差异.选取TaARF14基因起始密码子上游2 000 bp的序列进行启动子分析,见图 3.
由图 3可知,TaARF14-1A,TaARF14-1B和TaARF14-1D的启动子区域含有大量与植物正常生长发育(包括光响应、分生组织表达和低温响应)或植物激素(包括脱落酸、赤霉素和生长素响应)相关的顺式作用元件.3个基因的启动子区域都具有脱落酸和赤霉素响应的顺式作用元件,推测TaARF14-1A,TaARF14-1B和TaARF14-1D对小麦的生长发育具有重要作用,且TaARF14基因对赤霉素和脱落酸的响应模式具有较高的相似性,然而,只有TaARF14-1B和TaARF14-1D具有与生长素响应相关的顺式作用元件.进一步分析发现TaARF14-1A的启动子区域包括大量的“N”,是导致TaARF14-1A未检测到与生长素响应相关顺式作用元件的主要原因.
-
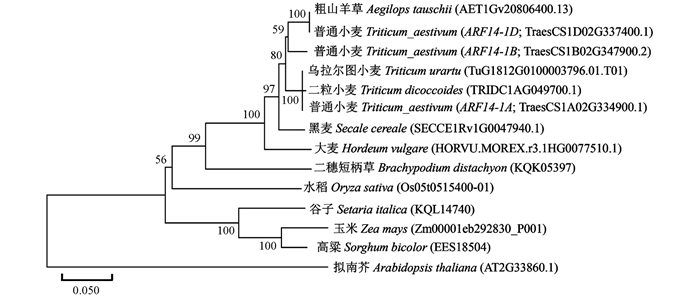
利用ARF14(TraesCS1A02G334900.1,TraesCS1B02G347900.2和TraesCS1D02G337400.1)的氨基酸序列在植物基因组数据中进行序列比对,构建了关于ARF14蛋白序列的系统发育进化树(图 4).14条氨基酸序列聚类为3个大类,其中拟南芥(Arabidopsis thaliana)单独聚类;玉米(Zea mays)、高粱(Sorghum bicolor)和谷子(Setaria italica)等聚为一个亚类;其他的麦类作物,包括普通小麦(Triticum aestivum)、乌拉尔图小麦(Triticum urartu)、二粒小麦(Triticum dicoccoides)、粗山羊草(Aegilops tauschii)、大麦(Hordeum vulgare)、二穗短柄草(Brachypodium distachyon)、黑麦(Secale cereale)等聚为一个亚类,表明ARF14基因在麦类作物中的系统发育进化具有较高的保守性.
-
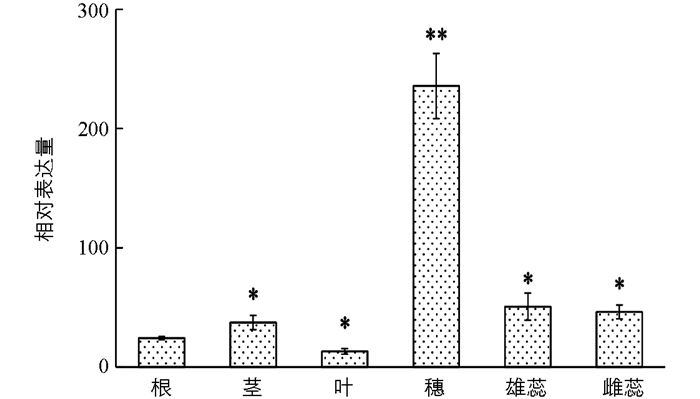
以小麦品种‘尚农6’的6个主要器官(包括根、茎、叶、穗、雄蕊和雌蕊)为模板,进行TaARF14基因的表达模式分析(图 5).结果表明:TaARF14具有明显的组织特异性表达,在小麦幼穗分化期(雌雄蕊分化期)的幼穗中具有极高的表达水平.此外,相对于小麦根部,TaARF14在抽穗期的雄蕊、雌蕊中也具有较高的表达水平,表明TaARF14基因在小麦的穗发育进程中,特别是花器官的发育(包括雄蕊和雌蕊)中具有极其重要的作用.
-
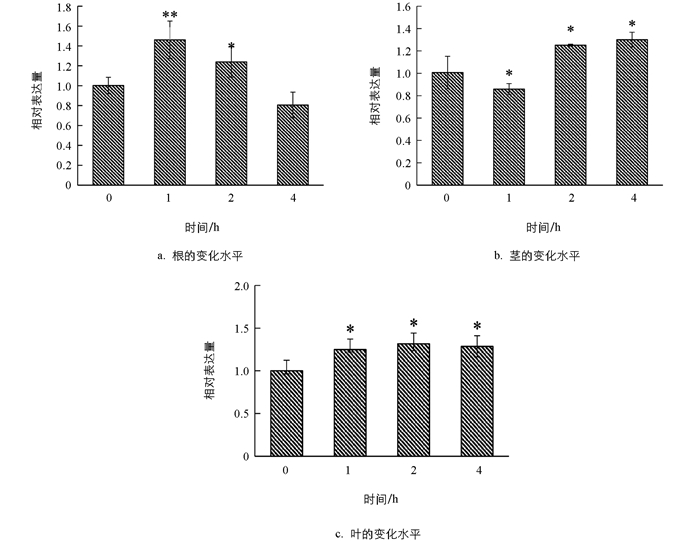
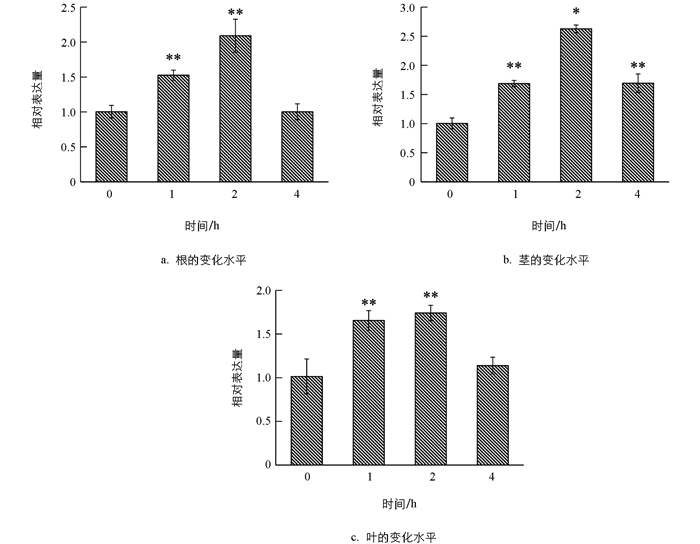
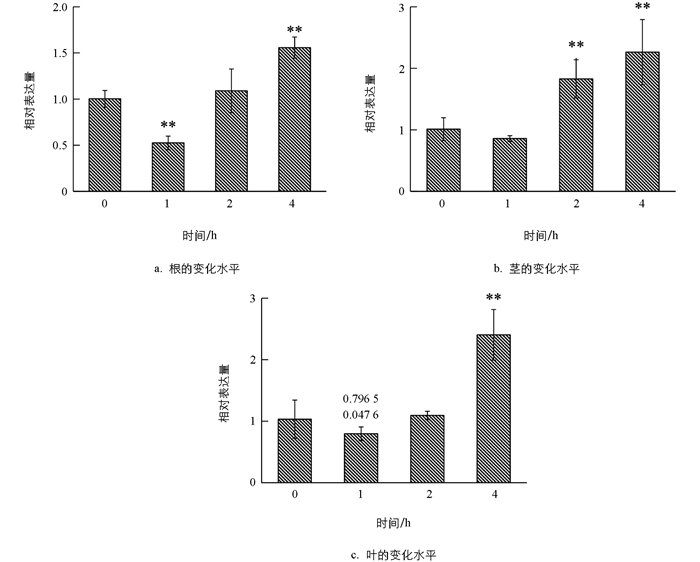
在施用IAA,GA和ABA后,利用qRT-PCR技术对TaARF14基因在小麦根、茎、叶中的表达模式进行分析,结果表明,在外源IAA的处理下(图 6),小麦根器官在1 h时基因表达水平呈显著上升,随后基因表达水平逐渐下降;在茎和叶中1 h时基因表达水平波动较小,随后基因表达水平趋于平稳.在外源GA的处理下(图 7),TaARF14在根、茎、叶中,基因表达变化趋势相似,总体表现为先上升后下降.在外源ABA的处理下(图 8),TaARF14基因的表达在根、茎、叶中总体表现为先下降后上升的趋势.整体来看,在3种外源激素的处理下,TaARF14基因表达水平在根、茎、叶中均具有明显的响应.
2.1. TaARF14基因的克隆
2.2. TaARF14基因的结构分析
2.3. TaARF14蛋白的系统发育进化树分析
2.4. TaARF14基因在不同组织和器官间的表达模式分析
2.5. TaARF14基因在激素胁迫下的表达模式
-
ARF基因家族的成员一般都具有相似的外显子/内含子结构,这为研究TaARF的进化关系提供了思路[22].相关数据表明,基因结构的相似性决定了其功能的相似性,具有相似结构的ARF基因可能具有相似的进化历史和功能[23].例如,在拟南芥中,AtARF8基因的抑制和过表达都会影响下胚轴的伸长和根的生长[24],通过转基因试验表明ARF8可促进或抑制拟南芥侧根的形成[25].类似地,TaARF4也已经被证实,其在决定小麦的根长和株高方面发挥着决定性的作用[26].这些结果表明来自不同植物物种的同源ARF基因可能具有相似的功能.大多数ARF基因具有不同的组织特异性表达模式,表明它们在不同组织/器官发育中的功能具有特异性. ARF17在拟南芥中通过直接调控CalS5基因的表达,从而影响花粉壁的形成[27].此外,OsARF19在花器官发育和植物结构中发挥着关键作用,具有明显的组织特异性表达[28].本研究中,TaARF14基因在小麦的穗发育进程中,特别是花器官的发育(包括雄蕊和雌蕊)中也具有明显的组织特异性表达,反映了其在小麦基因功能研究上的重要性.
植物激素几乎参与了陆地植物的所有发育过程,激素反应是植物发育和可塑性生长的基础.大量证据表明,施用外源激素(包括IAA,ABA和GA)会对小麦的生长发育和基因表达产生明显的影响,植物在胁迫响应方面具有其独特的响应机制[29].不同的ARF基因可能在植物发育过程中建立了多种独特的激素响应模式,如苋菜幼苗在不同浓度IAA和GA3的处理下,amaARF6基因均受调控,且生长素处理后基因表达水平发生了变化[30],推测TaARF14基因对植物的生长发育的调控方式在不同物种间存在差异.本研究中,TaARF14基因在响应激素胁迫方面表现出不同的表达模式,一般来说,特定的启动子控制基因在器官或组织中的表达[31],这个结果与TaARF14的3个同源基因(TaARF14-1A,TaARF14-1B和TaARF14-1D)的启动子区域分布着不同数量的与激素响应相关的顺式作用元件相关.本研究对TaARF14的3个同源基因进行了研究,发现TaARF14-1A,TaARF14-1B和TaARF14-1D的核酸序列和蛋白序列在结构上高度相似,且启动子区域存在大量与生长发育和激素调控相关的顺式作用元件,暗示了TaARF14基因功能的多样性,在施用外源IAA,ABA和GA处理后,qRT-PCR的结果也证实了这一推论.
综上所述,本研究在前人研究的基础上,对小麦转录因子TaARF14基因进行了克隆和表达分析,qRT-PCR表明,施用IAA,GA和ABA后,TaARF14基因的根、茎、叶等植物组织中对激素处理均具有明显的响应变化.本研究为后续TaARF14基因在激素胁迫下的基因表达和功能挖掘提供了重要的理论依据和试验依据.