



 下载:
下载:



-
开放科学(资源服务)标识码(OSID):

-
近年来,随着全球气候环境变化的加剧,植物在正常的生长发育过程中遭受干旱、低温、盐害等环境胁迫的现象时常发生,引起细胞内发生一系列不可控的生理生化变化,如活性氧积累、生物膜透性增大、渗透调节失调、光合速率下降等[1-2],从而影响植物的生长和产量.
多胺(polyamines,PAs)作为植物体内广泛存在的一类生理活性调节物质,主要以腐胺、亚精胺和精胺形式存在[3],在调节植物生长发育和缓解逆境胁迫等方面发挥重要作用[4-6]. S-腺苷甲硫氨酸脱羧酶(S-adenosylmethionine decarboxylase,SAMDC)作为多胺合成途径中一个关键限速酶,其主要功能是催化S-腺苷甲硫氨酸发生脱羧反应并提供丙氨基,分别在亚精胺合成酶和精氨合成酶的作用下实现腐胺向亚精胺和精胺的转化[7]. 目前已在甜橙[8]、苹果[9-10]、葡萄[11]、杜梨[12]、番茄[13]、辣椒[14]、甜瓜[15]、胡萝卜[16]、白菜[17]、康乃馨[18]等多种园艺植物中分离克隆出SAMDC基因,限速酶对不同的逆境胁迫会作出不同的响应,如胡萝卜SAMDC基因在高温、低温及盐胁迫等逆境胁迫中高表达,可能存在对多种逆境的交叉适应[16];橡胶树在干旱、盐害胁迫下其HbSAMDC1基因被诱导表达,但在冷害胁迫下却被抑制表达[19],表明SAMDC基因在植物抗旱、抗冷等逆境胁迫中起重要作用. 植物通过上调SAMDC基因的表达水平,以提高体内多胺的含量而增强植物对逆境胁迫的抵抗力. 例如烟草中过表达丹参SAMDC基因增强其抗旱性[20];在拟南芥中过表达甜瓜CmSAMDC增强了转基因拟南芥的耐盐性[21];车轴草TrSAMDC1受盐胁迫和干旱的诱导表达,超表达拟南芥后,植株体内多胺含量升高,增强了转基因植株的耐盐性和抗旱性[22].
桃(Prunus persica L.)作为我国一种重要的经济果树[23],其生长和产量易受到不良环境的影响,因此,挖掘利用抗逆基因资源及培育桃抗逆新品种是应对逆境胁迫最经济有效的途径之一. 目前已从桃基因组上克隆出一个多胺合成途径基因(PpADC)[24],转入番茄后内源Put和Spm含量明显增加,导致植株矮化和晚花表型[25],但未见多胺合成途径中其他相关基因的报道. 因此,本研究利用同源序列法从桃基因组上鉴定多胺合成途径中的关键基因(PpSAMDCs),对其进行克隆,并利用生物信息学手段进行进化分析、理化性质分析、蛋白质二级和三级结构的预测、信号肽、跨膜区和互作蛋白的预测,分析启动子元件并探究它们在非生物胁迫下的表达模式,旨在为探究桃PpSAMDCs基因的生物学功能奠定基础,也为桃的抗性育种提供重要的基因资源.
全文HTML
-
本研究所用桃来自于河南省新乡市卫辉蟠桃园,将层积处理过的实生毛桃种子播种于基质(体积比为草炭土∶蛭石=2∶1),所有植物材料在培养室温度25 ℃,16 h光照/8 h黑暗,相对湿度为60%的环境下生长. 选择长势一致、健壮的一个月龄的盆栽毛桃实生苗为试验材料,分别利用自然干旱、4 ℃低温和100 mM NaCl模拟自然环境下的逆境胁迫,每个处理3盆苗,每盆种植3棵,共9棵苗. 干旱处理在3 d和6 d时取样,低温处理在2 h和4 h时取样,盐胁迫在2 d和4 d时取样,取样时每棵植株剪取3个叶片,以未经任何处理的植株为对照. 取样后用锡箔纸包裹,置于液氮中速冻,放入-80 ℃超低温冰箱中保存、备用.
-
通过拟南芥TARI数据库(
http://www.arabidopsis.org/ )下载拟南芥5个AtSAMDCs基因的CDS序列,利用同源性搜索的方法,在Phytozome v13(http://phytozome.jgi.doe.gov/pz/portal.html# )网站选择桃基因组数据库(Prunus persica v2.1)进行Blast,鉴定出桃PpSAMDCs同源基因. 从网站下载PpSAMDCs基因的转录本序列、CDS序列、氨基酸序列及起始密码子ATG上游2 000 bp的启动子序列,以便用于基因克隆、生物信息学分析及表达分析等. -
采用总RNA提取试剂盒(生工生物工程股份有限公司,上海)提取桃各个样品总RNA,利用1%琼脂糖凝胶电泳和紫外分光光度计定性和定量检测RNA,采用PrimeScriptTM RT reagent Kit with gDNA Eraser(TaKaRa,大连)对检测完整的RNA反转录成单链cDNA,具体操作方法按照说明书进行.
-
依据获得的PpSAMDCs转录本序列,利用Primer premier 5软件设计每个基因的特异性全长引物. 引物由昆泰锐(武汉)生物技术有限责任公司合成,全长扩增引物序列如表 1所示. 以毛桃叶片cDNA为模板,利用Phanta Max Super-Fidelity DNA Polymerase(诺唯赞生物科技股份有限公司,南京)进行PCR扩增,扩增体系为50 μL,具体操作按照说明书进行. 扩增程序:95 ℃预变性5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s,循环35次;72 ℃延伸5 min. 取5 μL扩增产物进行琼脂糖凝胶电泳检测,利用DNA纯化试剂盒(SIMGEN,杭州)回收目的片段,连接pBLUNT载体(全式金生物技术有限公司,北京),并转化大肠杆菌DH5α,对阳性单克隆送至昆泰锐(武汉)生物技术有限责任公司进行测序.
-
利用MEGA4软件对PpSAMDCs蛋白和其他物种的SAMDC同源蛋白序列进行多序列比对,采用Neighbor-Joining法构建系统发育树,bootstrap值设置为1 000次重复. 利用NCBI网站中BLASTP程序对两个物种SAMDC蛋白序列进行同源性对比分析,采用DNAMAN软件进行作图.
-
对测序正确的PpSAMDCs基因编码的蛋白序列进行生物信息学分析,利用NCBI网站中CDD工具预测保守结构域,采用ExPASy网站中的ProParam工具预测PpSAMDCs蛋白的理化特性,运用SOPMA和SWISS-MODEL工具分别预测PpSAMDCs蛋白的二级结构和三级结构,利用LocSignaIP 4.1软件预测信号肽,利用TMHMM预测PpSAMDCs蛋白的跨膜结构域,利用STRING工具预测其互作蛋白,使用在线工具PlantCARE分析PpSAMDCs启动子上的顺式作用元件.
-
采用Primer premier 5软件设计PpSAMDCs基因的特异性定量引物(表 1),利用TB Green© Premix Ex TaqTM荧光定量试剂(TaKaRa,大连)分析逆境胁迫下PpSAMDCs基因的表达模式,具体的反应体系和反应程序参考说明书进行. 以桃PpTEF2为内参基因[26],对照中基因的表达归化为1,基因的相对表达量用FC=2-ΔΔCT公式[27]计算,每个样品进行3次生物学重复,数值用3次生物学重复的平均值±标准差表示.
-
使用Excel 2010对试验数据进行初步分析,利用T-test进行两个样本的差异显著性分析,然后采用GraphPad Prism软件制作图表.
1.1. 试验材料及处理
1.2. 桃基因组上PpSAMDCs基因的鉴定
1.3. 总RNA提取与cDNA合成
1.4. 序列扩增及测序
1.5. 同源性及进化分析
1.6. 生物信息学分析
1.7. 荧光定量PCR
1.8. 数据分析
-
参考下载的拟南芥AtSAMDC1(AT3G02470),AtSAMDC2(AT5G15950),AtSAMDC3(AT3G25570),AtSAMDC4(AT5G18930)和AtSAMDC5(AT3G17715)基因的CDS序列,采用同源搜索的方法在桃基因组数据库中进行比对,共鉴定出3个SAMDC同源基因. 运用NCBI网站中的CDD工具对这3个基因进行功能结构域的预测,结果显示都含有登录号cl03253,都隶属于S-腺苷甲硫氨酸脱羧酶家族. 根据它们在基因组上的位置分别命名为PpSAMDC1(Prupe.1G147600),PpSAMDC2(Prupe.1G299500)和PpSAMDC3(Prupe.3G243900).
以毛桃叶片cDNA为模板,利用扩增引物分别对PpSAMDC1,PpSAMDC2和PpSAMDC3进行PCR扩增,结果显示,分别获得一条清晰的特异性条带(图 1),与目的基因大小一致. 纯化回收后连接pBLUNT载体并转化大肠杆菌,挑选单克隆进行测序,测序结果显示,PpSAMDC1,PpSAMDC2和PpSAMDC3基因cDNA长度分别为1 158 bp,1 358 bp和1 197 bp,开放阅读框的长度分别为1 068 bp,1 074 bp和1 125 bp,分别编码355个、357个和374个氨基酸.
-
为了分析桃PpSAMDCs蛋白的进化关系,本研究以人HsSAMDC蛋白为外类群,选取桃和拟南芥等13种植物SAMDC蛋白[12]构建系统进化树,并进行同源性比对分析(图 2). 结果表明:桃PpSAMDC1蛋白处于整个植物进化的外群中,与其他物种亲缘关系都比较远,序列同源性都很低,在34.42%和43.59%之间. PpSAMDC2与樱桃PpSAMDC蛋白亲缘关系最近,同源性为97%,其次是苹果MdSAMDC1和白梨PbSAMDC3,同源性分别为88.57%和87.39%. PpSAMDC3与苹果MdSAMDC2亲缘关系最近,同源性最高,为89.6%. 上述几个物种均属于蔷薇科植物,说明桃PpSAMDC2和PpSAMDC3蛋白进化与植物进化具有一致性,而PpSAMDC1氨基酸进化程度比较低.
-
利用ExPASy网站中的ProtParam数据库分析PpSAMDCs蛋白的理化特性,结果显示PpSAMDC1,PpSAMDC2和PpSAMDC3蛋白相对分子质量分别为39.74 kDa,39.38 kDa和41.01 kDa,脂溶指数分别为72.76,80.03和81.12,等电点分别为5.10,4.94和5.12,亲水性系数均为负值;PpSAMDC1蛋白的不稳定指数小于40,而PpSAMDC2和PpSAMDC3蛋白不稳定指数大于40,表明PpSAMDC1蛋白是一个亲水的、稳定的酸性蛋白,而PpSAMDC2和PpSAMDC3蛋白为亲水的、不稳定的酸性蛋白.
-
利用SOPMA工具预测PpSAMDCs蛋白的二级结构,结果显示PpSAMDC1蛋白含有α螺旋32.68%、延伸链16.62%、β转角5.62%、无规则卷曲45.07%;PpSAMDC2蛋白含有α螺旋34.45%、延伸链18.77%、β转角8.40%、无规则卷曲38.38%;PpSAMDC3蛋白含有α螺旋33.96%、延伸链16.31%、β转角5.61%、无规则卷曲44.12%. 利用SWISS-MODEL在线工具对PpSAMDCs蛋白进行同源建模并预测三级结构,从中筛选GMQE(Global Model Quality Estimation)接近1、QMEAN(Qualitative Model Energy Analysis)分值接近0且序列比对相似度高大于30%的蛋白为模板,预测结果显示PpSAMDC1蛋白与山荆子SAMDC蛋白相似性最高,为85.55%,因此以该蛋白为模板进行PpSAMDC1蛋白的三级结构预测;PpSAMDC2蛋白与曼陀罗SAMDC蛋白相似性最高,为73.39%,因此采用曼陀罗SAMDC蛋白为模板构建PpSAMDC2蛋白的三级结构;PpSAMDC3蛋白与樱桃SAMDC蛋白相似性最高,为97.86%,用樱桃SAMDC为模板构建PpSAMDC3蛋白的三级结构. 利用LocSignalP 4.1工具预测PpSAMDCs蛋白信号肽,预测结果表明PpSAMDCs蛋白均无信号肽,属于非分泌性蛋白质. 利用TMHMM 2.0在线工具预测PpSAMDCs蛋白的跨膜结构,预测结果显示桃PpSAMDCs蛋白均无明显的跨膜结构,表明PpSAMDCs蛋白并非生物膜上的功能蛋白(图 3).
-
为揭示桃PpSAMDCs蛋白的功能及调控机制,利用STRING在线分析工具预测PpSAMDC1,PpSAMDC2和PpSAMDC3蛋白的互作网络. 预测结果表明:PpSAMDCs蛋白均与3个亚精胺合成酶(EMJ23702、EMJ19352、EMJ11235)、1个精胺合成酶(EMJ23466)、2个DNA(5-胞嘧啶)甲基转移酶(EMJ04405和EMJ09271)和3个S-腺苷甲硫氨酸合成酶(EMJ03317、EMJ08533、EMJ20481)发生互作. 此外,PpSAMDC1蛋白还与另一个精胺合成酶(EMJ23523)发生互作,PpSAMDC2和PpSAMDC3蛋白与另一个S-腺苷甲硫氨酸合成酶(EMJ24212)存在互作关系. 同时PpSAMDC1,PpSAMDC2和PpSAMDC3存在两两互作的关系(图 4).
-
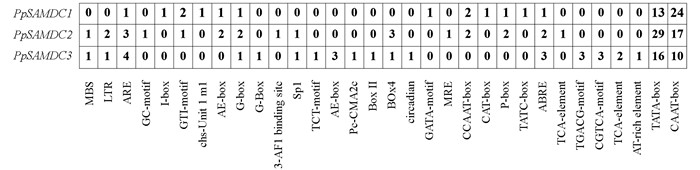
通过Plant CARE在线软件对PpSAMDCs启动子的顺式作用元件进行分析,结果显示:PpSAMDC2和PpSAMDC3均含有1个MYB参与的干旱诱导元件(MBS),分别含有2个和1个参与低温的诱导元件(LTR). PpSAMDC1,PpSAMDC2和PpSAMDC3都含有对厌氧诱导相关顺式作用元件(ARE)、多种光响应作用元件(I-box、GT1-motif、chs-Unit 1 m1、AE-box、G-box、G-Box、3-AF1 binding site、Sp1、TCT-motif、AE-box、Pc-CMA2c、Box Ⅱ、Box 4、circadian、GATA-motif)和脱落酸响应元件(ABRE),此外,PpSAMDCs启动子包含多个转录起始核心元件(TATA-box)、多个增强子区保守作用元件(CAAT-box)和1个分生组织表达相关调控元件(CAT-box). 序列分析结果表明,PpSAMDC1,PpSAMDC2和PpSAMDC3可能参与干旱、低温、光信号响应、厌氧调节等非生物胁迫及激素调控与分生组织发育等生物代谢过程(图 5).
-
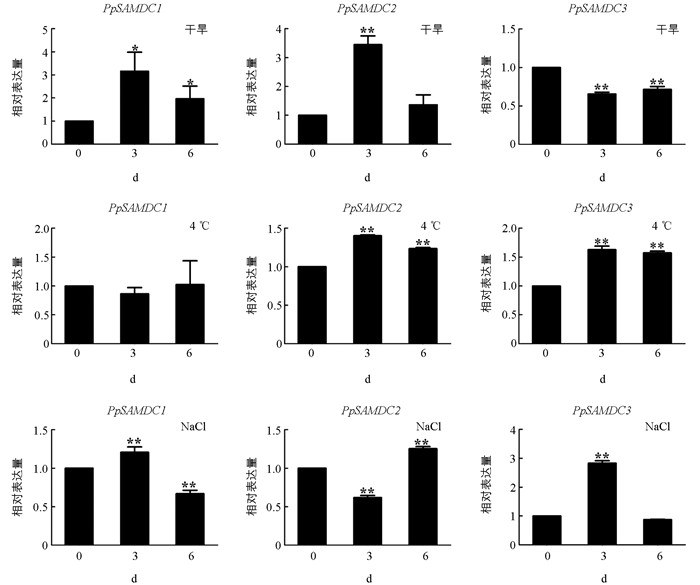
为了进一步探究桃PpSAMDCs基因对非生物胁迫的响应,本研究利用实时荧光定量PCR分析PpSAMDCs基因在干旱、低温和盐胁迫下的表达模式(图 6). 在干旱胁迫下,PpSAMDC1和PpSAMDC2基因的表达呈现先升高后下降的趋势,处理3 h表达量达到峰值,分别为对照(0 d)的3.15倍和3.44倍;干旱胁迫降低PpSAMDC3基因的表达,与对照相比,在3 d和6 d表达量分别下调35%和29%. 在低温胁迫下,PpSAMDC1的表达量无明显变化,但低温却诱导PpSAMDC2和PpSAMDC3基因的表达. 与对照(0 h)相比,在低温处理3 h和6 h时,PpSAMDC2基因表达分别上调40%和23%,PpSAMDC3基因的表达量分别上调62%和57%,差异均具有统计学意义. 盐胁迫促进PpSAMDC1和PpSAMDC3基因的表达,随着处理时间的延长均呈现先升后降的表达趋势,在盐处理2 d时达到最高值,分别为对照(0 d)的1.2倍和2.8倍;PpSAMDC2基因在盐胁迫下呈现先降后升的表达趋势,与对照相比,显著均具有统计学意义.
2.1. 桃PpSAMDCs基因的鉴定和克隆
2.2. 桃PpSAMDCs蛋白的进化分析及同源性分析
2.3. 桃PpSAMDCs蛋白的理化特性分析
2.4. 桃PpSAMDCs的蛋白结构、信号肽及跨膜结构分析
2.5. 桃PpSAMDCs互作蛋白预测
2.6. PpSAMDCs启动子顺式作用元件分析
2.7. PpSAMDCs基因在不同非生物胁迫下的表达分析
-
多胺是植物体内具有重要生理效应的生理活性物质,SAMDC是多胺合成途径中的一个关键酶,为精胺和亚精胺的生物合成提供丙氨基,在调控植物的生长发育和提高植物抗逆性方面起着非常重要的作用[28]. SAMDC基因已经在多种植物中克隆并进行功能验证,但未见关于桃SAMDC基因的报道. 鉴于此,本研究利用同源序列法从毛桃叶片cDNA中克隆出3个SAMDC同源基因,即PpSAMDC1,PpSAMDC2和PpSAMDC3,编码区长度分别为1 068 bp,1 074 bp和1 125 bp,分别编码355个、357个和374个氨基酸. 在其他植物中也发现存在多个SAMDC基因,比如拟南芥5个[29]、苹果8个[10]、棉花14个[30]、康乃馨2个[18]等,但有些植物基因组上只有单个SAMDC基因,比如柑橘[8, 31]、葡萄[11]、杜梨[12]、甜瓜[15]等. 一般来讲,同一家族的基因可能存在功能上的分化,有些基因出现功能的冗余,这些冗余基因在某些情况下可以增加生物体的生存能力,例如在环境变化或生物体发育过程中,它们可以提供额外的保护和适应能力[32].
利用生物信息学手段分析基因编码蛋白的结构有助于了解该基因的功能. 通过分析植物SAMDC同源基因的进化关系,PpSAMDC1氨基酸进化程度低,位于整个植物界的外群,而PpSAMDC2和PpSAMDC3与其他蔷薇科植物SAMDC蛋白亲缘关系近,与植物的进化较一致. 通过对桃PpSAMDCs蛋白序列的分析可知,桃PpSAMDCs蛋白均为亲水的酸性蛋白,二级结构以无规则卷曲和α螺旋为主,均无跨膜结构和信号肽,属于非分泌性蛋白. 这些蛋白结构的分析结果与砂糖橘CrSAMDC[31]、花生AhSAMDC[33]、刚毛柽柳ThSAMDC[34]、白菜BrSAMDC[17]等分析结果一致.
目前关于SAMDC的研究只集中在合成途径和功能研究上,而调控机制却知之甚少. Peng等[35]发现羊草中一个转录因子——脱水反应结合蛋白LcDREB2与LcSAMDC2启动上的DRE元件结合,增强了LcSAMDC2基因的表达,从而加强羊草的抗盐性. 本研究利用STRING工具分析PpSAMDCs与亚精胺合成酶、精胺合成酶、S-腺苷甲硫氨酸合成酶发生互作,表明参与多胺生物合成的基因存在互作关系. 此外,桃PpSAMDCs还与两个DNA甲基化转移酶发生互作,暗示PpSAMDCs可能参与到桃的表观遗传调控途径中,拓展了桃PpSAMDCs基因的功能. 互作网络的分析为进一步探索PpSAMDCs基因在多胺缓解逆境胁迫响应的调控机制提供了一定的理论支撑.
在逆境胁迫条件下,植物体内SAMDC基因表达量升高,通过增加多胺含量增强植物对逆境的抵抗能力,缓解逆境对植物造成的伤害[28]. Liu等[11]在葡萄中克隆出VvSAMDC片段,并且受盐胁迫的诱导. Zhao等[9]发现苹果MdSAMDC2在低温、盐和干旱胁迫后被诱导表达,表明该基因在苹果逆境胁迫中发挥重要调节作用. 本研究发现桃PpSAMDCs启动子上含有对干旱、低温、ABA激素等响应的顺式作用元件,定量PCR结果表明,桃PpSAMDCs基因对不同的非生物胁迫的应答反应不同,存在功能上的分化. PpSAMDC1基因对干旱及盐胁迫能快速响应,但对低温无应答;PpSAMDC2基因在干旱、低温和盐害等逆境胁迫下表达量均升高,功能更为广泛;低温和盐胁迫促进PpSAMDC3基因的表达,但干旱胁迫却抑制该基因的表达. 桃树相比其他果树耐旱性更强,推测桃抗旱强的原因可能是通过PpSAMDC1和PpSAMDC2合成多胺缓解干旱对植物的伤害. 苹果中的5个MdSAMDCs对逆境胁迫也存在不同的应答,但是MdSAMDC1能同时被5种逆境诱导,因此MdSAMDC1被认为是苹果SAMDC活性的主效基因[10]. 由于低温、干旱和盐胁迫都诱导PpSAMDC2基因的表达,所以PpSAMDC2基因是桃SAMDC基因家族中行使抗性功能的主要基因. 该研究挖掘和鉴定了桃抗逆的重要基因资源,为桃的抗逆新品种的定向培育奠定了理论基础.
-
本研究从桃基因组中鉴定出3个PpSAMDCs成员,并对其进化关系、理化特性、二级结构和三级结构、信号肽、跨膜区和互作蛋白进行分析,同时探究它们在干旱、低温和盐胁迫下的表达模式. 结果表明:这3个成员在不同逆境胁迫中可能扮演不同的角色,为探究桃PpSAMDCs基因的生物学功能和调控机制奠定基础,为培育桃抗逆品种提供重要基因资源.