


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
枯落物作为生态系统的重要组成部分,是土壤特有的发生层,具有增强水源涵养、调节水文过程、优化生态系统服务功能的效应[1-2]。枯落物层作为生态系统水源涵养功能的第二活动层,主要由植物茎、叶、花、果实、树皮及动物残体组成,能够截留降雨,减缓和抑制地表径流形成,防止土壤溅蚀,减少土壤水分蒸发,提高土壤储水能力以及净化水质[3-4]。在全球气候变化背景下,极端降雨和干旱事件频发,系统研究不同植物群落枯落物和土壤层水源涵养功能的差异,对于区域生态系统的可持续发展具有重要意义。
目前,国内外关于枯落物和土壤层的水源涵养功能开展了大量研究,并取得了丰硕成果。大量研究结果表明,植被枯落物和土壤层的水源涵养能力与植被类型、林龄、林分结构、枯落物分解程度以及枯落物蓄积量等密切相关[5-7]。但受气候条件、土壤母质以及地形地貌等因素的影响,同一植物群落在不同气候和地形条件下枯落物的分解速率和蓄积量存在明显差异,导致其枯落物和土壤层的水源涵养功能也存在显著差异。因为枯落物分解速率和蓄积量不仅直接影响枯落物层的水源涵养功能,还会通过影响土壤结构间接影响土壤层的水源涵养功能[6, 8-9]。因此,不同植物群落枯落物和土壤层的水源涵养功能存在明显的时空异质性。
三峡库区作为我国典型的生态脆弱区和水土流失重点治理区,近年来受城市建设和农业产业结构调整等的影响,库区土地利用格局发生了显著变化[10]。关于库区不同生态系统的水源涵养功能已有部分研究,如王鹏程等[9]通过对三峡库区10种主要森林植被类型枯落物蓄积量和持水能力的研究发现,不同森林类型枯落物蓄积量和持水能力存在较大差异;张焜等[11]对四面山的4种天然林枯落物的研究发现,阔叶林枯落物对降水的拦蓄效果最佳,而针阔混交林最差。现有研究主要集中在森林生态系统枯落物层的水源涵养功能,而关于土壤层水源涵养功能的研究相对较少。此外,研究区也主要集中在离库区较远的山区,而关于库区沿岸不同植物群落水源涵养功能的研究也相对较少。库区沿岸作为土地利用格局变化最为剧烈的区域,其水源涵养能力的变化会直接影响库区的生态环境。基于此,本研究在三峡库区腹地重庆市忠县的涂井乡,选取库区沿岸典型植物群落,通过野外采样和室内试验相结合的方法,系统分析不同植物群落枯落物持水能力和土壤层的储水能力,明确不同植物群落水源涵养功能的差异,以期为库区沿岸土地利用结构的优化调整及土地资源的可持续发展提供理论依据和实践指导,助力三峡库区的绿色可持续发展战略的实施。
全文HTML
-
研究区(108.10°E,30.30°N)位于重庆市忠县涂井乡友谊村的秦岭小流域内,海拔为120~800 m,毗邻长江干流,属典型的山地丘陵峡谷地带区。气候类型为典型的湿润季风气候,多年平均降水量约为1 100 mm,多年平均温度为19 ℃,降雨主要集中在雨季4-10月,约占全年降水总量的70%。研究区土壤为紫色土,由紫色砂泥岩发育而成,土层较薄,具有入渗性能好、抗蚀性能差的特点。近年来受人类活动的影响,区域内土地利用结构发生了显著变化,集中体现在经果林大面积增加,使得区域水土流失和面源污染加剧。目前流域内主要以林地、果园、灌木地、草地以及坡耕地为主。
-
2023年8月,在对小流域全面野外探察的基础上,选取了流域内坡度、坡向、海拔以及植被盖度相近的柏树(Cupressus funebris)、马尾松(Pinus massoniana)、桉树(Eucalyptus)、柑橘(Citrus reticulata)、灌木、草地和坡耕地7个典型植物群落,其中柏树、马尾松、桉树、柑橘种植密度通过在样地内调查3个10 m×10 m的样方计算得到,玉米种植密度通过在样地内调查3个2 m×2 m的样方计算得到。各样地基本信息如表 1所示。
-
采样时间选在采样前一周无降水的情况下进行,在选定样地内,采用“S”型方法设置3个0.5 m×0.5 m的样方框,用游标卡尺分别测定枯落物未分解层、半分解层以及枯落物层总厚度(每个样点重复测量10次)。测量完成后,将样方框内所有未分解和半分解枯落物分别装入自封袋,带回实验室。枯落物中的泥沙用清水冲洗后,装在档案袋中,在85 ℃条件下烘干至恒质量后称质量,分别记录未分解和半分解枯落物的蓄积量。枯落物收集过程中,通常将颜色未发生明显变化、保持原有形态、无分解痕迹的枯落物归为未分解枯落物;将无完整叶轮廓、颜色已发生明显变化、叶片已分解破碎的枯落物归为半分解枯落物。同时现场用自封袋收集少部分未分解和半分解枯落物,带回实验室后立即称质量,得到枯落物湿质量(m1),然后将枯落物转移到信封中,在85 ℃条件下烘干至恒质量后称质量,得到枯落物干质量(m2),枯落物自然含水率(R0)的计算公式如下:
测定完枯落物蓄积量后,称取一定量的干枯落物放入尼龙网袋中,在清水中分别浸泡1/12、1/4、1/2、1、2、4、6、10、24 h后取出晾干至无水滴滴落时迅速称质量,然后计算不同浸泡时间下枯落物的持水量、吸水速率及拦蓄量等指标[12]。通常,随着枯落物浸泡时间的增长,枯落物持水量会趋于稳定,此时的持水量被认为是枯落物的最大持水量,其对应的拦蓄量即为枯落物最大拦蓄量,相关指标计算公式如下所示[6]:
式中:R0和Rm分别为枯落物自然含水率和最大持水率(%);We和Wm分别为枯落物有效拦蓄量和最大拦蓄量(t/hm2);M为枯落物蓄积量(t/hm2);Mi为持续浸水i时间的枯落物重量(g);Md为浸水试验所用枯落物干质量(g);t为枯落物持续浸水时间(h);Vi为持续浸水i时间枯落物吸水速率[×103 g/(kg·h)]。
-
土壤水文效应指标主要包括土壤容重、饱和含水量、田间持水量、总孔隙度、毛管孔隙度以及有效蓄水量,其中土壤饱和含水量、田间持水量、总孔隙度以及毛管孔隙度采用环刀法测定,土壤容重采用烘干称质量法测量[13]。基于测定的土壤水文指标进一步计算了土壤有效蓄水量[14],其计算公式为:
式中:S为土壤有效蓄水量(t/hm2);h为土壤层厚度(m);TP为总孔隙度(%);CP为毛管孔隙度(%);ρ水为水密度(×103 kg/m3)。
-
不同植物群落枯落物及土壤层水文效应指标的差异性采用单因素方差分析(one-way ANOVO)中的多重比较法(LSD)进行分析;枯落物持水量和吸水速率随浸水时间的关系采用非线性回归的方法进行分析,所有数据均使用SPSS 21.0软件处理,图形绘制采用Origin Pro 2021软件完成。
1.1. 研究区概况
1.2. 样地选择
1.3. 枯落物蓄积特征调查及持水能力测定
1.4. 土壤水文效应指标测定
1.5. 数据处理
-
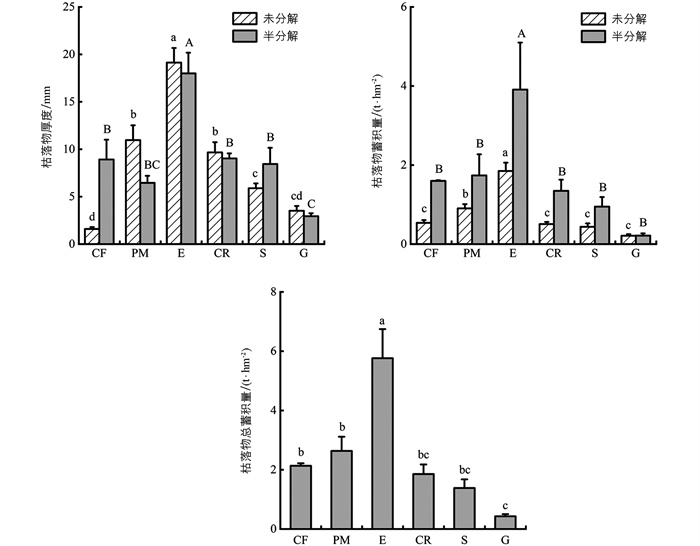
不同植物群落枯落物厚度和蓄积量存在一定差异。各植物群落未分解层和半分解层的枯落物厚度分别为1.60~19.13 mm和2.94~18.00 mm,其中,桉树林未分解层和半分解层枯落物厚度均为最大,而未分解层和半分解层枯落物厚度最小的分别为柏树和草地。不同植物群落未分解、半分解以及总的枯落物蓄积量分别为0.21~1.85、0.22~3.91、0.43~5.76 t/hm2,且未分解、半分解以及总的枯落物蓄积量最小的均为草地,最大的均为桉树(图 1)。未分解和半分解枯落物蓄积量占枯落物总蓄积量的比例为25.16%~49.61%,均小于50%。总体来看,未分解和半分解枯落物蓄积量从大到小均依次为:林地、柑橘、灌木、草地。
单因素方差分析结果表明,桉树未分解和半分解层枯落物厚度均显著大于其他植物群落(p<0.05);而柏树未分解层和草地半分解层枯落物厚度均显著小于其他植物群落(p<0.05)。对于枯落物蓄积量,桉树未分解、半分解和总枯落物蓄积量均显著大于其他植物群落;而草地未分解枯落物蓄积量仅显著小于桉树和马尾松(p<0.05),总蓄积量与其他植物群落(除柑橘和灌木外)差异均有统计学意义。
-
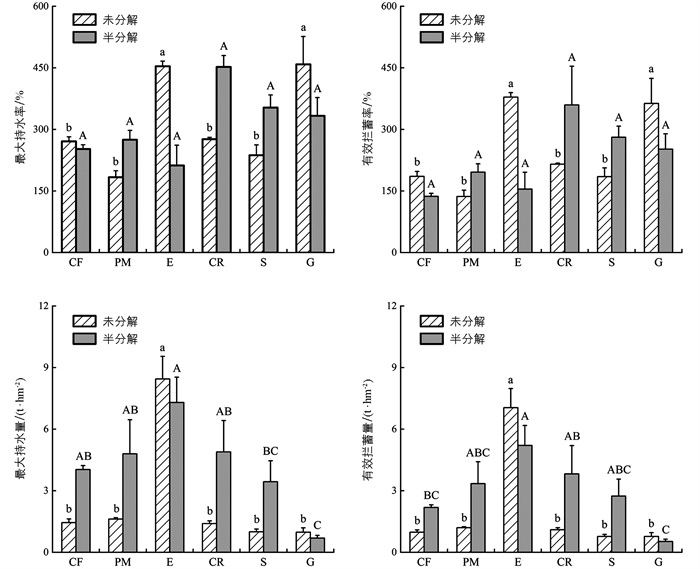
不同植物群落枯落物持水能力差异有统计学意义。各植物群落未分解枯落物最大持水率、有效拦蓄率、最大持水量和有效拦蓄量分别为183.82%~458.74%、136.70%~378.76%、0.98~8.45 t/hm2和0.78~7.05 t/hm2,半分解枯落物则分别为212.45%~452.38%、136.86%~359.75%、0.69~7.30 t/hm2和0.53~5.21 t/hm2。不同植物群落未分解和半分解枯落物的最大持水率和有效拦蓄率没有明显的变化规律,但桉树枯落物的最大持水量和有效拦蓄量均为最大,而草地为最小;且桉树和草地未分解枯落物的最大持水量和有效拦蓄量均大于半分解枯落物,而其他植物群落则与此相反(图 2)。总体来看,三峡库岸不同植物群落枯落物对降雨的有效拦蓄量表现为桉树为最大,然后依次为柑橘、马尾松、柏树、灌木地和草地。
单因素方差分析结果表明,桉树和草地未分解枯落物最大持水率和有效拦蓄率均显著大于其他植物群落(p<0.05),而柑橘半分解枯落物最大持水率和有效拦蓄率均大于其他植物群落,但差异无统计学意义。此外,桉树未分解枯落物的最大持水量和有效拦蓄量均显著大于其他植物群落(p<0.05),而草地半分解枯落物最大持水量和有效拦蓄量均显著小于其他植物群落(p<0.05)。
-
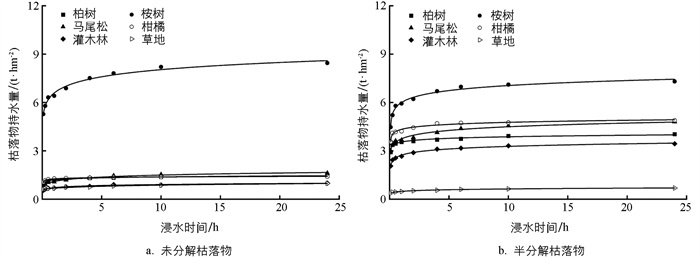
不同植物群落未分解和半分解枯落物具有相似的持水过程。总体表现为先迅速增加(0~1 h),后缓慢增加(1~10 h),最后趋于稳定的变化规律(>10 h)(图 3)。回归分析结果表明,枯落物持水量随时间的变化可以用Q=aln(t)+b的对数函数进行拟合,拟合方程的R2均大于0.9,表明拟合效果很好(表 2)。
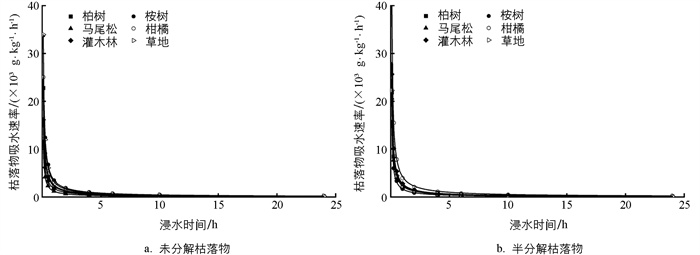
不同植物群落枯落物吸水速率存在一定差异,其中未分解枯落物吸水速率从大到小依次为:草地、桉树、柑橘、柏树、灌木、马尾松,而半分解枯落物吸水速率从大到小依次为:柑橘、灌木、草地、桉树、柏树、马尾松,其中马尾松未分解和半分解枯落物的吸水速率均为最小。而不同植物群落枯落物吸水速率随浸水时间的增加表现为先迅速减小(0~1 h),后缓慢减小(1~10 h),最后趋于稳定的变化规律(>10 h)(图 4)。回归分析结果表明,不同植物群落枯落物吸水速率随时间的变化可以用V=atb的幂函数进行拟合,拟合方程的R2均大于0.99,表明拟合效果很好(表 3)。
-
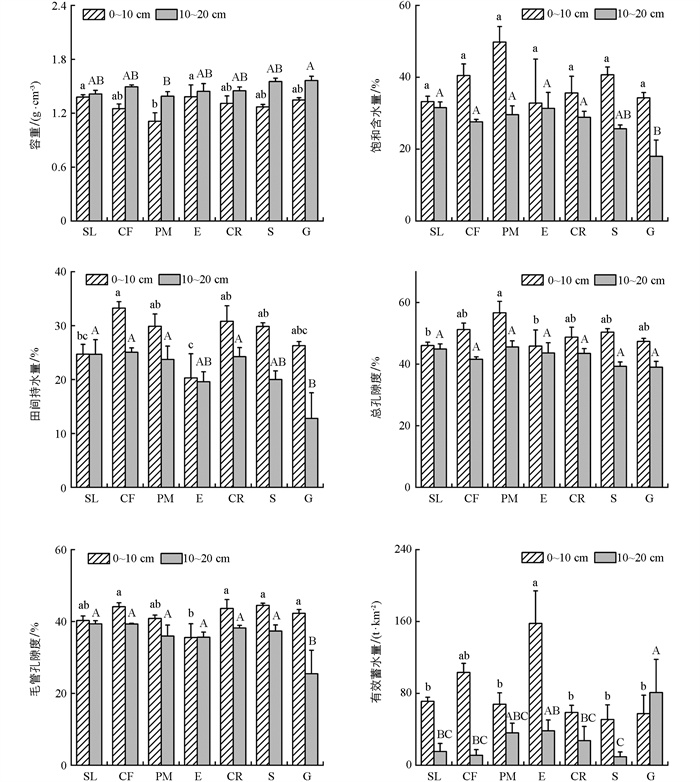
不同植物群落土壤水文性质存在一定差异。各植物群落0~10 cm和10~20 cm土层土壤容重、饱和含水量、田间持水量、总孔隙度、毛管孔隙度和有效蓄水量的范围分别为:1.27~1.38 g/cm3、32.80%~49.81%、20.31%~33.25%、45.87%~56.65%、35.53%~44.49%、50.99~157.87 mm和1.14~1.56 g/cm3、17.99%~31.55%、12.78%~25.06%、39.02%~45.59%、25.48%~39.35%、9.39~81.04 mm(图 5)。其中,桉树0~10 cm土层的土壤容重和有效蓄水量最大,但饱和含水量、田间持水量、总孔隙度和毛管孔隙度最小;10~20 cm土层则表现为草地的土壤容重和有效蓄水量最大,而饱和含水量、田间持水量、总孔隙度和毛管孔隙度最小。单因素方差分析结果显示,桉树0~10 cm土层土壤有效蓄水量显著大于其他植物群落(柏树除外),而草地10~20 cm土层土壤有效蓄水量显著大于其他植物群落(桉树和马尾松除外)。各植物群落土壤容重均随着土层深度的增加而增加,而土壤饱和含水量、田间持水量、总孔隙度、毛管孔隙度和有效蓄水量(草地除外)则随土层深度的增加而减少。
-
三峡库区不同植物群落枯落物和土壤层(0~20 cm)的有效蓄水量为63.90~208.42 t/hm2,其中桉树的有效蓄水量最大,然后依次为草地、柏树、马尾松、柑橘、坡耕地和灌木。通过对比各植物群落枯落物层和土壤层水源涵养能力的贡献发现,枯落物层水源涵养能力的贡献为0~5.88%,而土壤层(0~20 cm)的贡献介于94.12%~100.00%,表明土壤层的水源涵养能力决定了各植物群落水源涵养能力的大小,其贡献率在94%以上(表 4)。
2.1. 不同植物群落枯落物蓄积特征
2.2. 不同植物群落枯落物层水文效应
2.2.1. 枯落物持水能力
2.2.2. 枯落物持水过程分析
2.3. 不同植物群落土壤层水文效应
2.4. 不同植物群落水源涵养能力
-
受植被类型、冠层盖度和人类活动等多因素的影响,不同植物群落的水热条件存在差异,进而影响枯落物的蓄积量及分解速率,导致枯落物的水源涵养功能发生变化。本研究发现三峡库区不同植物群落的未分解枯落物蓄积量均小于半分解枯落物蓄积量(图 1),这与西南地区的部分研究结果相一致[15-17]。主要原因在于枯落物的分解受降雨和温度等气候条件影响显著[3],而本研究区毗邻长江,夏季降雨充沛,且温度高,为微生物和土壤动物的生长和发育提供了适宜的环境,加速了枯落物的分解过程,从而导致未分解枯落物蓄积量小于半分解枯落物。
枯落物最大持水量和有效拦蓄量反映了枯落物的持水能力,是衡量枯落物潜在水源涵养能力的重要指标,其大小与枯落物蓄积量、初始含水量等密切相关[18-20]。通常,枯落物水源涵养能力随枯落物蓄积量的增加而增强[3]。本研究发现,桉树未分解和半分解枯落物的最大持水量和有效拦蓄量均为最大,而草地均为最小(图 2),这主要是因为桉树的枯落物蓄积量最大而草地最小(图 1)。此外,虽然桉树和草地未分解枯落物蓄积量小于半分解枯落物(图 1),但未分解枯落物最大持水量和有效拦蓄量均大于半分解枯落物(图 2),这主要与枯落物的持水率有关。因为枯落物水源涵养能力除了与枯落物蓄积量有关外,还与枯落物持水率有关,通常当枯落物自然含水量相差不大时,枯落物的最大持水率和有效拦蓄率越大,其最大持水量和有效拦蓄量越大[21]。而桉树和草地未分解层枯落物的最大持水率和有效拦蓄率均显著大于半分解层枯落物,使得桉树和草地未分解枯落物的最大持水量和有效拦蓄量均大于半分解层枯落物(图 2)。
不同植物群落未分解和半分解枯落物持水过程可以分为3个阶段,即快速吸水—缓慢吸水—趋于稳定,这与大多数研究结果相一致[3, 6]。枯落物的吸水速率主要受水势差控制,野外条件下枯落物较为干燥,水势较低,在降雨或泡水条件下,巨大的水势差使得枯落物快速吸水;而随着枯落物含水量的增加,水势差减小,导致吸水速率逐渐减小,直至趋于稳定[6]。因此,枯落物持水量和吸水速率随浸水时间的增加分别呈现“先迅速增加后缓慢增加至趋于稳定”和“先迅速减小后缓慢减小至趋于稳定”的变化规律,这一变化规律可以通过对数函数和幂函数很好地进行拟合(图 3、图 4),这与前人在不同地区的研究结果基本一致[3, 6, 22]。
-
土壤层作为生态系统水分调节和储存的媒介,其水源涵养能力与土层厚度、土壤容重以及孔隙特征等密切相关[23-26]。而土地利用变化引起植被枯落物蓄积量及根系特性等的改变,会驱动土壤容重和孔隙等发生变化,进而影响土壤的水源涵养功能。本研究发现,在库区不同植物群落中,0~10 cm土层土壤有效蓄水量最大的为桉树,10~20 cm土层最大的为草地,但桉树和草地在相应土层的总孔隙度和毛孔隙度均不是最大(图 5)。其主要原因是,土壤的水源涵养能力主要受控于土壤的非毛管孔隙度,通常土壤非毛管孔隙度越大,其水源涵养能力越强[27-28]。综合来看,库区0~20 cm土层桉树林的土壤水源涵养能力最大,然后依次为草地、柏树、马尾松、柑橘、坡耕地、灌木地。而灌木地土壤水源涵养能力为最小,这与前人研究结果不同。可能是因为该区域人地矛盾较为紧张[10, 29],灌木仅生长在土层较薄,无经果林和乔木林种植条件的荒地上,加之牛羊的踩踏,导致灌木地的土壤水源涵养能力较差。
综上可知,三峡库区的乔木林枯落物和土壤层均表现出较高的水源涵养能力;而草地虽然枯落物层的水源涵养能力较低,但其土壤层水源涵养能力较强,使其总体水源涵养能力仍较强(图 2、图 5、表 4)。因此,仅从水源涵养功能的角度考虑,乔木和草地是库区相对合理的植被模式,但在乔木的选择上,应综合考虑其耗水及其对土壤结构的长期影响。此外,柑橘和坡耕地的土壤水源涵养能力几乎相等,表明坡耕地建设成经果林后对土壤的水源涵养能力几乎没有影响,但其水源涵养能力总体较差。因此,在经果林的建设过程中可以通过在经果林下种植草本绿肥来改良土壤结构,增强土壤的水源涵养能力,以提高其适应极端干旱天气的能力。
相较于土壤层的水源涵养能力,库区各土地利用枯落物层的水源涵养能力较弱,贡献率均小于6%(表 4),这与大多数的研究结果一致[24, 30]。但现有研究多采用野外采样和室内分析相结合的方法,单独研究枯落物层和土壤层的潜在涵养水源能力,关于枯落物涵养水源的间接作用没有量化,而枯落物可以通过增加地表粗糙度减缓径流的汇流时间,促进入渗,进而增加土壤的蓄水量[31-32],因此,现有研究结果可能大大低估了枯落物在水源涵养中发挥的重要作用,未来研究需结合控制试验,进一步探讨枯落物与土壤层的协同作用,以全面评估其在水源涵养中的贡献。
-
本研究通过野外采样和室内分析相结合的方法,对三峡库区(库岸)典型植物群落枯落物蓄积特征、持水过程及枯落物和土壤层的水源涵养功能进行了系统研究,发现库区不同植物群落枯落物蓄积特征和持水能力均存在一定差异,其中枯落物蓄积量从大到小依次为林地(桉树、马尾松、柏树),果园,灌木地,草地,且未分解枯落物蓄积量均小于半分解枯落物;对于枯落物持水能力,桉树未分解枯落物最大持水量和有效拦蓄量显著大于其他植物群落,而草地半分解枯落物最大持水量和有效拦蓄量显著小于其他植物群落,且各植物群落(除桉树和草地外)未分解枯落物的最大持水量和有效拦蓄量均小于半分解枯落物。此外,不同植物群落未分解和半分解枯落物的持水量和吸水速率随浸水时间的增加分别呈对数函数增加和幂函数减小的变化趋势。不同植物群落土壤层(0~20 cm)有效蓄水量也存在差异,表现为桉树最大而灌木最小。综合来看,三峡库区不同植物群落枯落物和土壤层水源涵养功能从高到低依次为:桉树、草地、柏树、马尾松、柑橘、坡耕地、灌木,且各植物群落水源涵养能力的大小均受控于土壤层的水源涵养能力。仅从水源涵养功能的角度来看,乔木和草地是库区尤其是库岸比较合理的植被模式。