


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
硼作为植物生长发育所必需的微量营养元素,参与并维持细胞壁的合成及其结构的稳定,同时还参与绿色植物的光合作用和糖的转运等多种代谢过程[1]。目前,我国约有3.3×105 km2土地存在缺硼问题,尤其在降水充沛的东南地区,土壤缺硼现象尤为突出。硼的缺乏严重制约了当地作物的优质高产。与此同时,植物对硼的需求具有严格的适应范围,极易出现硼毒害或硼缺乏症状[2]。适量施用硼肥能够有效提高植物的品质和产量[3]。Xu等[4]研究了不同硼含量施用方法对番茄果实品质和风味的影响。研究发现,无论是根施还是叶面喷施,均能提升番茄品质,且在叶面喷施的情况下,番茄红素含量随着硼含量的增加而增加,表明硼对番茄红素的合成具有促进作用。
番茄红素是一种类胡萝卜素,研究表明其具有较强的抗氧化作用,能够增强人体免疫力,对预防和治疗前列腺疾病、心血管疾病及癌症具有一定的效果[5]。番茄红素的生物合成是在多种酶的催化作用下完成的,主要通过MEP和MVA两条合成途径。其合成过程本质上是叶绿素逐渐降解,并转化为以番茄红素和β-胡萝卜素为主的类胡萝卜素。目前,已有研究表明,番茄红素的关键合成基因及酶包括八氢番茄红素合成酶(PSY)[6]、八氢番茄红素去饱和酶(PDS)[7]、类胡萝卜素异构酶基因(ZISO和CRTISO)[8]、ζ-胡萝卜素脱氢酶(ZDS)[9],此外,还有与番茄红素合成相关的基因如LCYB等。洪敏[10]通过病毒诱导基因沉默技术干扰枇杷果实中PSY基因的表达,结果表明,沉默处理后的果实侵染部位呈绿色,而对照组果实为黄色,且在果肉中均能检测到TRV病毒分子,PSY基因的表达量降低了55.6%,总类胡萝卜素含量降低了46.8%,这表明PSY基因在枇杷果实类胡萝卜素积累中起着正向调控作用。Romero等[11]通过病毒诱导基因沉默技术(Virus-Induced Gene Silencing,VIGS)沉默PDS基因,发现沉默后的水果番茄红素含量显著降低。Fantini等[12]利用病毒诱导基因沉默结合高分辨率液相色谱技术研究了ZDS的功能,结果发现ZDS沉默后的水果类胡萝卜素含量增加,且番茄红素几乎完全消失。Efremov等[13]的研究表明,Z-ISO的表达与成熟番茄果实中类胡萝卜素的含量呈正相关,同时品种和野生物种的转录分析表明,Z-ISO的表达水平可能是决定成熟番茄果实中类胡萝卜素积累的关键因素。Ronen等[14]研究发现,随着番茄果实颜色逐渐加深,番茄红素合成的上游基因PSY的表达水平上升,而下游代谢基因LCYB的表达水平则不断降低,直至无法检测到。这些研究均表明,相关基因的表达与番茄果实中番茄红素的含量密切相关。
转录因子在调控植物生长发育和应激反应中起着重要作用,其中bZIP家族是植物中一类关键的转录因子家族。目前,已有研究成功鉴定了许多真核生物的bZIP基因家族成员。研究表明,番茄中含有69个bZIP基因成员,且这些基因成员可分为9个亚家族[15]。bZIP家族转录因子由DNA结合的碱性区和可变的亮氨酸拉链区组成,其中碱性区高度保守,由约20个氨基酸残基构成,包含一个核定位信号和一个保守的N-X7-R/K基序,后者能识别并结合启动子特定的核酸序列;亮氨酸拉链区的N末端可以与酸性结构域结合,形成同源或异源二聚体,从而实现转录的抑制或活化[16]。研究发现,bZIP转录因子在果实成熟方面也发挥着重要作用。有研究表明,番茄中bZIP转录因子SIHY5的敲除会降低花青素的积累。在光照条件下,番茄SIHY5能够激活花色苷生物合成基因的转录,从而调节花色苷的合成,并影响由光调节的果实着色过程[17]。在拟南芥的研究中发现,磷酸化作用会影响衰老bZIP转录因子GBF1与DNA的结合[18]。此外,有研究认为,拟南芥的C/S1 bZIP网络是协调植物生长和应激反应的一个信号枢纽[19]。Hsieh等[20]的研究发现,bZIP转录因子基因SlAREB在风干和高盐度条件下显著提高了番茄的抗逆性。这些研究表明,bZIP转录因子在不同物种中可能发挥相似或不同的作用。
番茄红素的含量与其品种有着密切关系,不同品种的番茄表现出不同的性状。例如,有些品种在成熟后呈现黄色[21],而有些则呈现红色甚至深红色。余越等[22]通过VIGS技术对SlWRKY53b基因进行沉默研究,发现该基因在抑制番茄果实成熟和降低番茄红素含量方面起着重要作用。目前,关于bZIP家族基因的研究主要集中在植物的非生物胁迫和生物胁迫响应上,而针对番茄果实成熟的研究相对较少,尤其是在番茄中,只有少数bZIP转录因子(TFs)被鉴定并进行了功能表征。
全文HTML
-
试验盆栽土壤全氮含量为0.725 g/kg,碱解氮含量为136.73 mg/kg,有效磷含量为53.13 mg/kg,速效钾含量为182.3 mg/kg,土壤有机质含量为20.78 g/kg,pH值(水/土=1/1)为5.88,有效硼含量为0.023 mg/kg。供试番茄品种分别为‘渝番713’和‘LBR1731’,由重庆市农科院蔬菜花卉所提供。试验在西南大学资源环境学院温室大棚内进行。大田试验在潼南科光种苗中心进行,番茄为盆栽供试品种。
-
盆栽时间为2022年2月至2022年6月,硼质量浓度设置参照前期试验结果[23],试验设置5个浓度的硼砂:0,0.5,1,2,4 mg/L,分别记作CK,A1,A2,A3,A4。取5 kg过10 mm筛网筛选的风干土,装入塑料盆中,混合加入N(150 mg/kg)、P(120 mg/kg)和K(180 mg/kg)肥料,其中K以硫酸钾(K2SO4)形式加入,P以磷酸二氢氨(NH4H2PO4)形式加入,N以NH4H2PO4和尿素的形式加入。70%作为底肥,剩余30%根据番茄长势酌情进行追肥(移栽21 d后第一次追肥)。土壤混合均匀并平衡2~3周后,每盆移栽2株长势一致、体型健壮的番茄幼苗。在番茄开花期,按照设计的处理方案进行喷洒,每个处理进行3次重复,随机排列。试验期间使用去离子水进行浇灌,盆土水分保持在田间持水量的70%左右。
VIGS试验于2023年1月至2023年6月在重庆市潼南区太柏路西的重庆科光种苗中心的大田中进行,供试品种为盆栽品种。硼质量浓度设置为0、1和2 mg/L,共3个处理,其中不施硼为对照组(CK)。每个处理设3次重复,共设计18个小区。大棚规格约为30 m×8 m,试验根据温室实际情况,选择其中3厢进行种植。每厢种植两行,约1/6厢为一个小区,每个小区种植20株番茄苗,保护行宽度及株行距根据当地种植习惯设置。所有处理的有机肥、氮磷钾肥施用量及病虫害防治均依据当地管理习惯。在番茄开花期,根据设计的处理进行喷洒,每个处理3次重复,随机排列。
-
选取番茄开花期第7~8位叶片用于总RNA提取、mRNA纯化、cDNA文库构建及高通量测序(由深圳华大基因股份有限公司进行),并进行3次重复测序。
为将RNA-seq读长(reads)与基因组进行比对,首先使用Trimmomatic v.0.39[26]和fastp软件对原始读数质量进行评估。过滤后的读数利用HISAT2 v2.1.0与番茄参考基因组(
https://solgenomics.net/ftp/genomes/Solanum_lycopersicum/Heinz1706/annotation/ITAG4.0_release/ )比对。然后,使用StringTie v1.3.3(Pertea et al.,2016)进行转录本组装,并使用内置的prepDE.py脚本计算读数。差异表达基因(DEGs)的筛选使用DESeq2 package v1.16.1(Michael et al.,2014),筛选条件为:表达倍数变化大于1.5,FDR(False Discovery Rate)<0.05。GO和KEGG背景注释信息分别通过www.geneontology.org 和www.genome.jp/kegg 网站获取,并通过clusterProfiler包进行富集分析。使用Blast2GO将番茄蛋白序列与Swissport数据库(https://www.expasy.org/resources/uniprotkb-swiss-prot )进行比对和注释,并通过超几何分布法确定差异基因富集的GO术语。为了进一步探讨与差异表达基因相关的信号通路,使用TBtools软件进行KEGG通路富集分析。转录组测序数据差异表达分析使用limma包,筛选阈值为p<0.05,log2FC>1或log2FC<-1。差异基因的火山图通过ggplot2包绘制,bZIP家族基因的表达水平热图通过pheatmap包生成。基因表达水平的相关性通过Hmisc包计算,并使用corrplot包绘制相关性热图。根据p值与相关系数r筛选共表达基因,最终通过Cytoscape绘制共表达网络。 -
根据基因的保守序列选取靶片段,并使用primer3Plus进行实时荧光定量引物设计,随后在NCBI数据库中进行目的基因匹配验证(表 1、表 2)。
采用PowerUpTMSYBRTMGreen Master Mix(Applied Biosystems,Vilnius,Lithuania)试剂盒配制反应体系,并使用荧光定量PCR仪(QuantStudioTM 1 System,USA)进行定量PCR分析。PCR反应程序为:95 ℃预变性10 min,95 ℃变性30 s,60 ℃退火30 s,进行40个循环;随后从60 ℃升温至95 ℃,检测产物溶解曲线。所有试验均重复3次。
目标基因的相对表达量使用2-ΔΔCT方法计算,其中CT值表示荧光信号达到设定阈值所经历的反应循环数[27]。
-
使用primer3Plus设计带有酶切位点BamHI和SmaI的TRV同源臂引物,进行目的基因的扩增。通过限制性内切酶酶切载体,并进行同源重组构建重组载体。随后,将重组载体转化至大肠杆菌感受态DH5α中,并使用含有卡那霉素的抗性固体培养基进行筛选。通过菌检PCR获得阳性克隆后,提取质粒并送往北京擎科新业生物技术有限公司(武汉)进行测序确认。测序结果正确的克隆菌株进行扩繁,提取质粒,并采用冻融法转入农杆菌。农杆菌转化筛选使用含有卡那霉素、利福平和庆大霉素的培养基,挑选单一菌落进行菌液PCR,并用琼脂糖凝胶电泳检测进行验证[28]。确认正确的农杆菌液后,在3种抗性筛选(卡那霉素、利福平和庆大霉素)培养基上进行过夜震荡培养。次日,离心收集菌体,用等体积的缓冲液(10 mmol/L氯化镁、10 mmol/L MES、200 μmol/L乙酰丁香酮,pH=5.6)重悬菌体,并调节OD600为1.0左右。将pTRV1与转化农杆菌的培养液按1∶1比例混合,室温黑暗孵育3 h后[29],使用1 mL无菌注射器注射番茄果柄和相邻叶片[22]。注射后将番茄果实置于黑暗环境中培养24 h,随后恢复正常生长条件。接下来,在注射后的7 d内进行连续采样以检测沉默效率,以注射pTRV2-PDS的果实作为参照。在约3周后,待番茄果实出现明显症状(如与同期相比不变色或变色延迟)时,取样后用qRT-PCR来测定基因表达量,并测定番茄红素含量及其相关合成基因的表达水平。试验重复3次。
-
使用SPSS Statistics 26.0软件进行数据分析,具体方法包括主成分分析、单因素方差分析(One-way ANOVA)、相关性分析(Pearson相关系数法)。对于不同处理间的均值差异显著性,采用LSD检测法进行比较,显著性水平设定为p<0.05。
1.1. 试验材料
1.2. 试验方法
1.3. 测定指标与方法
1.3.1. 番茄红素及硼含量
1.3.2. 转录组测序
1.3.3. bZIPs基因表达
1.3.4. 病毒介导的番茄SlbZIP36的沉默
1.4. 数据处理与统计分析
-
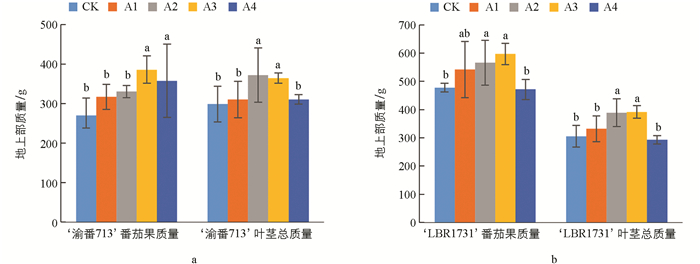
如图 1所示,随着供硼水平的增加,番茄植株的茎叶总质量与果实质量呈现先增加后减少的趋势。与对照组相比,两个品种的番茄植株茎叶质量分别在A2和A3处理下达到最大值,分别为对照组的1.24倍和1.28倍,且与其他处理组相比具有显著差异。‘渝番713’的茎叶总质量在A2处理下开始有显著差异,在A4处理下略有下降,但仍高于对照组,总质量相比对照组增长了3.65%~24.35%。对于‘LBR1731’,随着供硼水平的增加,茎叶总质量也在A2处理下显著增加,但在A4处理下有所下降。除A4处理外,其茎叶总质量分别增长了8.66%~28.23%。总体而言,两个品种在各处理组间的差异较为显著。随着供硼水平的增加,两个品种的番茄果实总质量也呈现逐步增加的趋势。与对照组相比,两个品种的番茄均在A3处理下达到了最大值,分别为对照组的1.43倍和1.25倍。‘渝番713’果实质量随着供硼水平增加呈现先增加后减少的趋势,在A2处理下表现出显著差异,A4处理虽呈下降趋势,但仍高于对照组,果实质量增长了17.49%~43.05%。‘LBR1731’的变化趋势与‘渝番713’类似,在A2处理下出现差异,在A3处理下达到最大值,除A4处理外,果实质量分别增长了13.39%~24.94%。这些结果表明,适当的硼处理有助于促进番茄植株的生长。
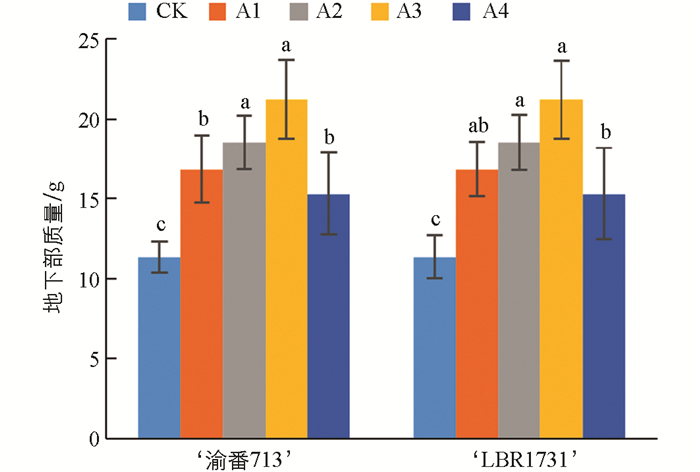
如图 2所示,两个番茄品种的地下部质量均随硼水平的增加而增加。与对照组相比,两个品种的地下部质量在A3处理下分别达到了最大值,分别为对照组的1.87倍和1.51倍,增长幅度为34.86%~87.23%和6.23%~51.62%。同时,图中还显示,与A3处理相比,番茄植株在A4处理下的地下部质量明显降低,但仍高于对照组。表明适宜的硼浓度有助于促进番茄植株的生长,而过量的硼供应可能会抑制植株的生长。
-
如表 3所示,随着供硼水平的增加,‘渝番713’的根部含硼量升高,在A2处理下达到最大值,相比对照组增长了32.20%,各处理间差异较为明显。与之相反,‘LBR1731’的根部含硼量随着供硼水平的增加而逐渐减少,在A2处理下达到最低值,但在A4处理下虽有所回升,仍低于对照组,且各处理间差异不显著。对于番茄茎部,两个品种的茎含硼量均随供硼水平增加而增加。‘渝番713’在A4处理下的茎含硼量达到最大值,为对照组的1.44倍,各处理间差异不显著;‘LBR1731’的茎含硼量在A2处理下达到最大值,为对照组的1.12倍,不同处理间差异不显著。
叶片是番茄植株中含硼量最高的部位,其硼含量接近根部、茎部和果实硼含量的总和。‘渝番713’的叶片含硼量随着供硼水平的增加而增加,在A4处理下达到最大值,较对照组增加了11.33%~45.55%,不同处理间存在明显差异;‘LBR1731’的叶片含硼量在A4处理下达到最大值,为对照组的1.33倍,较对照组增加了7.63%~33.24%,不同处理间差异显著。不同供硼处理下,两个品种的番茄果实硼含量差异不显著。‘渝番713’的果实硼含量随供硼水平的增加呈上升趋势,在A3处理下达到最大值,为对照组的1.30倍,增幅为19.49%~29.66%。‘LBR1731’的果实硼含量在A3处理下也达到最大值,为对照组的1.20倍,增幅为3.36%~20.17%。
-
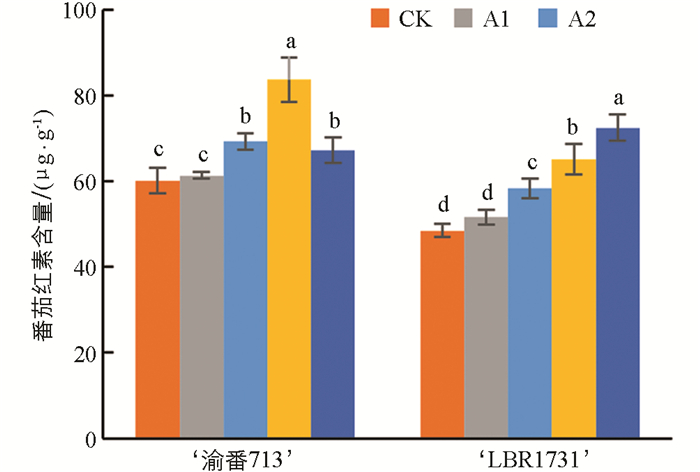
如图 3所示,‘渝番713’的番茄红素含量随着供硼水平的增加而增加,在A3处理下达到最大值,是对照组的1.73倍。虽然在A4处理下略有下降,但仍高于对照组,且不同处理间差异显著。‘LBR1731’的番茄红素含量随着供硼水平的增加呈现线性增长,在A4处理下达到最大值,各处理的番茄红素含量相比对照组分别增加了6.43%~49.46%,是对照组的1.06~1.50倍。上述结果表明,硼含量对番茄红素含量有显著影响,能够有效提升番茄的品质。
-
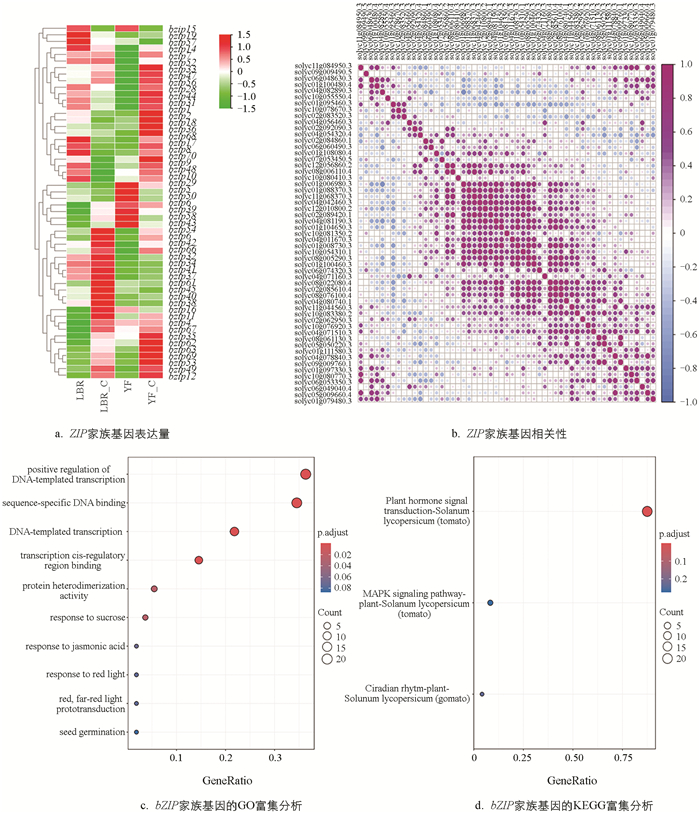
通过检测果实中bZIP家族基因的表达量,筛选出60个差异表达的基因(图 4a)。在两种番茄果实中,大部分基因的表达呈上调趋势。尤其是bZIP7、bZIP49和bZIP4等基因在未处理的条件下表达量较高,而bZIP49在施硼处理后表达量显著下降。对施硼处理前后55个bZIP家族基因(去除表达量为0的基因)在‘LBR1731’与‘渝番713’中的相关性进行分析(图 4b),发现bZIP49与bZIP36、bZIP1、bZIP7等10个基因具有高度相关性。同时,bZIP家族基因的GO和KEGG富集分析显示,这些基因主要与蔗糖反应、茉莉酸反应以及植物激素信号转导等生物过程密切相关(图 4c、4d)。
-
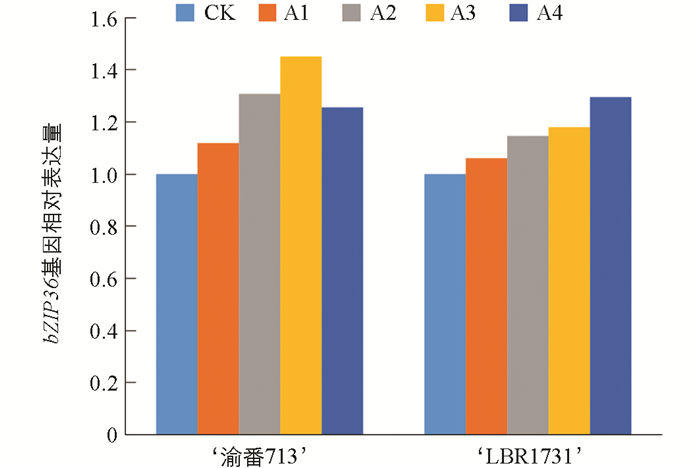
由图 5所示,两个品种番茄果实中bZIP36基因的相对表达量均随着供硼水平增加而增加,分别在A3处理(2 mg/L)和A4处理(4 mg/L)下达到最大,分别为对照的1.45倍和1.29倍,说明硼在一定范围内可以增加番茄果实中bZIP36基因的相对表达量。各处理间表达量差异显著。
-
为了研究bZIP相关转录因子的表达量与番茄果实中硼含量的相关性,进行了Pearson相关性分析。由表 4可知,番茄果实中硼含量与bZIP36呈显著正相关(r=0.660,p<0.05),说明bZIP36与番茄果实硼含量高度相关。
-
由表 5可知,bZIP36与bZIP49呈现负相关,bZIP36与番茄红素含量呈现显著正相关(r=0.930,p<0.05)。根据此结果可以推测,供硼水平增加促进了bZIP36的表达,而bZIP36的表达进一步促进了番茄红素的合成。
-
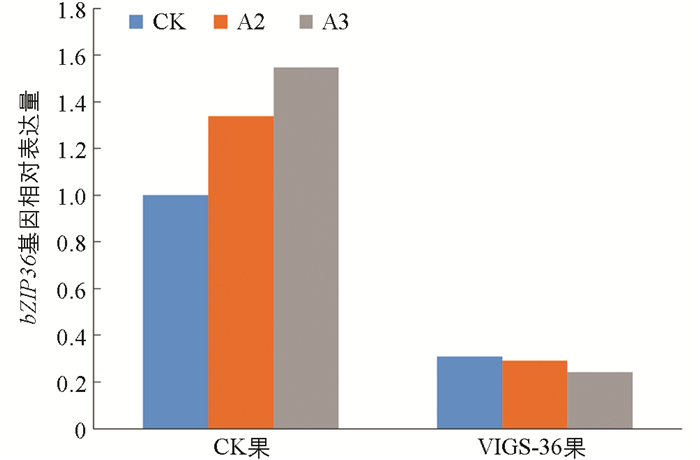
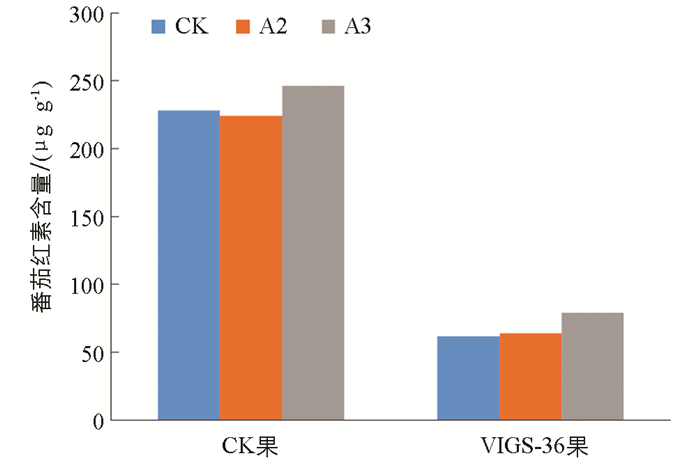
图 6所示为不同硼处理下VIGS产生的效果,在硼处理下,果实颜色有变深趋势,且可以明显看出沉默SlbZIP36基因对番茄果实颜色变化有明显的抑制作用。由图 7所示,随着供硼水平的增加,A2和A3处理下普通果实内bZIP36表达量逐渐增加,分别为对照组的1.34倍和1.55倍,说明硼元素可以促进bZIP36基因的表达。但3种处理下注射了pTRV2-bZIP36病毒载体的果实内bZIP36表达量变化却相反,随着施硼量增加,病毒果实内bZIP36表达量呈降低趋势,CK处理下表达最高为0.31,整体沉默效率在70%以上。由图 8所示,与CK相比较,A3处理下番茄红素含量增加,比对照增加了17.1 μg/g。同时注射了pTRV2-bZIP36的病毒果实与对照组相比,其番茄红素含量远低于对照组,说明沉默bZIP36会导致番茄果实番茄红素含量下降,成熟延迟。
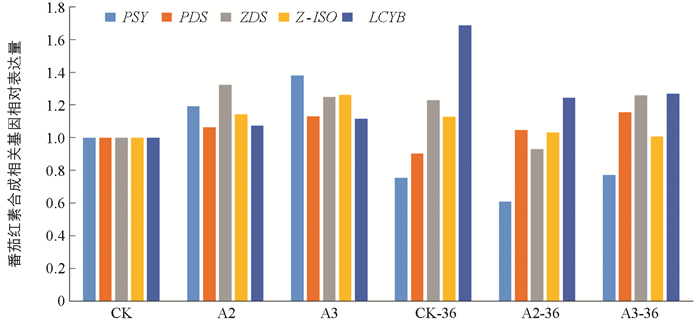
由图 9所示,与CK相比较,PSY相对表达量随着施硼含量增加而呈现上调表达趋势。PDS相对表达量和PSY变化趋势一致,在A3处理下达到最大值1.12。ZDS相对表达量随着施硼含量的增加,表达量下降到0.93,随后表达量上调,上调后表达量为对照组的1.26倍。Z-ISO相对表达量在不同施硼处理下略微上调随后下调与CK一致,变化幅度很小整体几乎没有变化。LCYB(番茄红素β-环化酶基因)相对表达量随供硼水平增加上调表达,最大表达量为1.26。当以普通果实CK为对照,3个处理下注射了病毒载体果实内PSY相对表达量呈现下调表达,为对照组的0.61~0.77倍。PDS相对表达量随供硼量增加而上调表达,但在CK处理下病毒果实内PDS表达量低于对照组,说明沉默bZIP36会下调PSY和PDS的表达。ZDS相对表达量在A2处理下低于对照组,在其余两个处理上均上调表达。Z-ISO相对表达量随供硼水平增加呈现下调趋势,但总体高于对照组,在CK处理下果实内的表达量最高。LCYB相对表达量为CK处理下的1.69倍,在之后的处理下表达量下调。说明沉默bZIP36导致了番茄红素合成基因PSY的下调表达以及LCYB的上调表达。
如表 6所示,SlbZIP36与PSY呈极显著相关(r=0.984,p<0.01),PSY(八氢番茄红素合酶)是类胡萝卜素生物合成途径上的第一个合成酶,说明bZIP36的表达会促进PSY的上调表达,促进番茄红素的合成。SlbZIP36与PDS呈正相关(r=0.272),但是与ZDS、Z-ISO、LCYB呈负相关(r=-0.121、r=-0.416、r=-0.399)。LCYB可以催化番茄红素转化合成β-胡萝卜素,SlbZIP36表达量越高,LCYB表达越低,转化为胡萝卜素的番茄红素越少,番茄红素含量越高。
2.1. 不同供硼水平对番茄植株生物量的影响
2.2. 不同供硼水平对番茄硼量的影响
2.3. 不同供硼水平对果实番茄红素含量的影响
2.4. 硼诱导下bZIP家族转录因子转录组研究分析
2.5. 不同硼水平下番茄果实bZIP36基因表达量分析
2.6. bZIPs基因表达量与番茄果实硼含量的相关性分析
2.7. bZIP36基因表达量与番茄果实番茄红素含量的相关性分析
2.8. SlbZIP36沉默分析
-
硼是植物生长发育所必需的微量元素,适量的硼能够提高植物的产量和质量。番茄的果实质量和产量是衡量外界调控因子对植株生长影响的重要指标之一[30]。本试验通过外源施加不同质量浓度的硼,发现‘渝番713’和‘LBR1731’两个品种的番茄果实质量随着供硼水平增加呈现上升趋势,但在A4处理下,两品种的果实质量均有所下降。表明当供硼水平为2 mg/L时,番茄生长条件达到最优,而高质量浓度硼(A4处理)可能抑制果实的生长。因此,在适宜的硼质量浓度范围内,果实生长与硼质量浓度呈显著正相关。这与李梅兰等[31]的研究结果一致,他们发现,在一定质量浓度范围内,施用硼肥显著提高了番茄果实质量;同时,卢一铭等[23]的研究也证实了不同质量浓度的叶面施硼肥对番茄单果质量的正向影响,进一步验证了本试验的结果。在番茄植株的茎叶总质量和地下部质量方面,变化趋势与果实质量类似,都在2 mg/L供硼水平下达到最大值,表明这一质量浓度范围最有利于番茄植株的生长。徐龙水[32]的研究也发现,番茄在不同硼质量浓度条件下的生长发育受到不同程度的影响,进一步支持了本试验的结果。
近年来,基因转录组测序技术的快速发展为植物基因转录组研究提供了更加先进的平台。李金昊等[33]通过分析番茄SULTR基因家族的转录谱,发现其启动子区域包含多种参与植物激素及逆境胁迫响应的顺式作用元件。同时,脱落酸、乙烯、茉莉酸甲酯和水杨酸处理能够迅速诱导多个SlySULTR基因的表达,表明这些基因可能参与激素信号传导过程。毕晨曦等[34]通过对盐胁迫下小麦bHLH转录因子家族进行鉴定,利用差异表达基因的GO富集分析发现,这些基因在分子功能、生物过程和细胞组成等方面表现出显著差异。王春妹等[35]通过干旱胁迫下藜麦种子的糖代谢转录组研究,发现了54 000多个差异表达基因,并通过KEGG富集分析发现,糖代谢过程主要富集在淀粉和蔗糖代谢、半乳糖代谢、果糖和甘露糖代谢等3个通路中。该发现为进一步研究干旱胁迫下藜麦种子糖代谢的分子调控机制提供了重要参考。上述转录组分析为研究样本提供了更深入的见解。因此,本研究设置了不同的硼质量浓度处理,以探讨不同硼水平下的差异表达基因、富集变化与相关性等。通过对两个番茄品种‘LBR1731’和‘渝番713’在不同施硼处理下的转录组数据进行分析,发现两个品种番茄的上调基因和下调基因趋势相同,但‘LBR1731’和‘渝番713’的上调基因数量低于下调基因数量。虽然两个品种的差异表达基因数量相近,但在某些情况下,两个品种之间的差异较大,这表明硼对番茄的影响可能与品种有关。对差异基因进行基因本体功能(GO)富集注释后,发现两个品种番茄共同的差异基因主要富集在细胞代谢过程中。进一步分析硼处理后两个品种间功能基因的差异,发现其差异主要集中在木葡聚糖代谢和细胞壁生物合成等过程,表明细胞在硼处理过程中作为信息传递者起着重要作用。此外,KEGG分析显示,氨基酸和生物激素代谢通路富集了较多的差异基因,提示硼处理可能影响番茄的抗病性、氨基酸合成及生长,进而可能对番茄的抗病性和口感品质产生一定影响。
在植物中,bZIP转录因子(TF)是最大且最具多样性的转录因子家族之一。研究表明,bZIP TF家族在不同生物体中的数量各不相同[16]。通过基因敲除或过表达等方法,广泛的研究表明,bZIP TF家族的成员参与植物许多器官和组织的分化、胚胎发生、种子成熟、花的开放与生长以及维管发育[36]。此外,bZIP TF家族还被证明是信号通路中的关键成分,参与植物对非生物和生物胁迫(如渗透、缺氧、干旱、高盐度、低温胁迫及病原体感染)的反应[37-40]。目前,番茄中的bZIP TF SlAREB1(脱落酸反应元件结合蛋白1)已得到广泛研究,发现它在果实成熟过程中对环境胁迫和代谢调控的响应中发挥着重要作用,并作为ABA信号转导与生物胁迫反应的连接点[40]。Pan等[41]的研究表明,干旱处理导致SlbZIP38表达量下调,且过表达SlbZIP38的转基因番茄植株比野生型抗旱性更差。
番茄红素合成相关代谢基因与番茄红素的合成密切相关,八氢番茄红素的合成是限速步骤,决定了类胡萝卜素生物合成的通量。因此,PSY是该途径的关键调节酶[42]。番茄红素β-环化酶通过环化线性分子番茄红素的两端,生成β-胡萝卜素,而LCYB通过一端与另一端的ε-环化酶环化,产生α-胡萝卜素。马超等[43]利用RNAi技术调控番茄中的LCYB基因,结果表明,LCYB基因的表达被抑制,转基因番茄果实中番茄红素含量显著增加。杨亮等[44]通过CRISPR/Cas9技术定向编辑滞绿基因SlSGR1,发现SlSGR1表达量显著降低,PSY1表达量显著提高,从而有效提升番茄果实中番茄红素、β-胡萝卜素等类胡萝卜素的含量。表明相关番茄红素关键合成酶基因的表达量变化与番茄红素含量密切相关。本研究通过病毒诱导基因沉默技术,在不同硼处理下发现,SlbZIP36的表达量随着供硼水平的增加而上调。同时,番茄果实内的番茄红素含量与SlbZIP36呈极显著正相关。注射pTRV2-bZIP36病毒载体的果实表现出明显的成熟延缓,同时期的普通果实已经转色至红色成熟期时,病毒果实仍保持在绿熟期。基因定量结果表明,沉默SlbZIP36后,果实内PSY的表达量降低,LCYB等基因的表达量增加。进一步分析发现,SlbZIP36与PSY呈极显著正相关,而与LCYB的表达量呈负相关。表明SlbZIP36参与番茄红素的合成过程,能够通过提高PSY的表达,起到正向促进作用。
-
随着供硼质量浓度的增加,番茄的生物量呈上升趋势,果实质量在A3处理下达到最大值,且番茄红素含量也随之增加。同时,随着施硼水平的提高,番茄红素合成关键基因(包括PSY、PDS、ZDS等)的表达量也在增加。SlbZIP36与番茄红素含量之间呈极显著正相关,同时与果实硼含量也呈显著正相关。对bZIP家族基因的GO和KEGG分析结果显示,该基因家族在蔗糖反应、茉莉酸反应以及植物激素信号转导等方面具有重要影响。沉默SlbZIP36显著抑制了PSY等基因的表达,从而导致番茄红素合成降低。相关性分析进一步发现,SlbZIP36与番茄红素含量呈显著正相关。