


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
昆虫在适应生存环境的过程中,进化出了多种不同类型的附肢[1],这些附肢的多样性最显著地体现在数量、形态和位置上,如头部体节分化成的口器、胸部体节分化成的翅膀和足以及腹部体节分化成的外生殖器等。对于完全变态的昆虫而言,其幼虫阶段的足可分为3种基本类型:①多足型,特点在于其胸部有3对胸足,同时腹部可以附着1至多对腹足;②寡足型,仅具有3对胸足,腹部不具备任何足;③无足型,幼虫的每个体节均没有足的结构。
从分子发育生物学的视角分析,昆虫附肢在数量、形态及位置上的差异所受的遗传调控机制主要源于进化过程中基因调控网络动态变化及功能分化。附肢的演化涉及多个基因表达网络通过层级调控的相互作用。Hox基因家族是一个重要的发育调控基因群,最早在果蝇中被发现[2],负责控制昆虫躯体的模式发育,其排列顺序决定了躯体不同体节的特征[3-4]。Hox基因对昆虫附肢的发育起着关键的调控作用,尤其是在附肢的发育部位和形态特征方面[5-7]。在不同昆虫物种中,Hox基因功能的分化被认为是导致各种附肢类型产生的重要原因[8-9]。作为转录调控因子,Hox基因需通过精确控制不同下游靶基因的表达来调节各类附肢的发育,因此鉴定和解析附肢发育中Hox基因的下游靶基因,对于理解昆虫附肢的分化具有重要的学术价值。家蚕作为鳞翅目昆虫,拥有丰富的遗传突变资源[10-12]。在西南大学家蚕遗传资源基因库中,有一类E群突变体,其特征是附肢的数量异常,表现为过多或缺失[13-15],其中EN和ENk突变体胚胎的腹足与胸足类似,而ECa和EMc突变体胚胎在腹部节段没有腹足,相比之下野生型Dazao胚胎在腹部节段有4对正常腹足,同时这4种突变体胚胎均有3对正常的胸足位于胸部节段,而在腹部相同体节表现出不同的附肢形态:①腹部有正常的4对腹足;②腹部有类似胸足的足;③腹部无足。利用E群突变体的形态学表型差异及差异基因表达谱,分析Hox基因及其下游靶基因调控家蚕附肢发育和特化的机制[16-20],可以解析家蚕附肢发育与形态特化的分子调控网络。
本研究以野生型Dazao的EN、ENk、ECa和EMc发育腹肢突起出现期的胚胎为实验材料进行转录组测序,通过比较转录组数据,筛选出参与家蚕附肢发育及负责胸足和腹足特化的重要相关基因。本研究不仅在分子水平上揭示了家蚕胸足与腹足表型的发育分化机制,更为解析昆虫附肢形态多样性的Hox基因调控网络提供了关键的实验依据,为阐明昆虫附肢发育可塑性及其演化发育机制奠定理论基础。
全文HTML
-
家蚕EN、ENk、ECa、EMc突变体及野生型Dazao均来自于西南大学家蚕遗传资源基因库,在温度25 ℃、相对湿度75%、12 h光照12 h黑暗条件下饲养。
将手术刀片、解剖针、玻璃培养皿用0.1% DEPC水完全浸泡12 h去除RNA酶,锡箔纸包裹后于160 ℃烘箱中高温灭菌4~6 h。准备一次性无RNA酶的研磨棒、1.5 mL离心管及均浆器备用。将发育至腹肢突起期的家蚕野生型Dazao的EN、ENk、ECa、EMc突变体胚胎自25 ℃培养箱中转移至操作台,使用灭菌刀片在蚕卵侧面切开小口,将其浸入盛有1×PBS缓冲液的玻璃培养皿中,于显微镜下用解剖针从开口卵壳中轻柔分离出胚胎,清除附着浆膜及卵黄残留物,用刀片切取胚胎腹部环节,使用1 mL移液枪将所取组织移到1.5 mL RNase-Free管中,每管包含20~30个胚胎的组织样,取3管做生物学重复,用液氮速冻,保存在-80 ℃冰箱中备用。
材料准备结束后立即用Omega公司的MicroElute®Total RNA Kit进行RNA提取。液氮研钵研磨,粉末混合TRK lysis Buffer裂解液和β-巯基乙醇,静置,离心,上清混合等体积75%乙醇,转移至离心柱,静置,离心,弃废液;依次用HiBind Buffer洗脱1遍,离心,RNA washing Buffer洗脱离心2遍,空离,晾干,加水静置离心。检测浓度后用Takara公司生产的PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time)进行反转录得到cDNA,检测浓度后稀释5倍于-80 ℃冰箱中保存备用。
根据基因的cDNA序列在NCBI上设计定量引物,BLAST对比其特异性。根据选择的定量引物,送擎科生物公司进行合成。进行实时荧光定量检测,检测体系:10 μL 2×NovoStart® SYBR qPCR SuperMix,0.2 μL正向引物,0.2 μL反向引物,1~2 μL Template,用RNase Free H2O补足至20 μL。将上述混合液按每个样品3个技术重复,分装至96孔PCR板中。离心后于荧光定量PCR仪中进行检测,步骤如下:95 ℃ 1 min;95 ℃ 20 s,60 ℃ 1 min采集荧光,40个循环。
-
RNA-seq文库在北京诺禾致源完成,通过磁珠富集mRNA,使用fragmentation buffer断裂mRNA,采用随机六碱基引物合成一链cDNA,再通过dNTPs和DNA聚合酶合成双链cDNA。纯化后进行末端修复、加A尾、连接测序接头,并选择特定大小的片段。使用PCR扩增构建cDNA文库,经过Qubit 2.0定量、Agilent 2100评估片段大小,Q-PCR精确量化,确保浓度大于2 nmol/L。Illumina测序后,使用Trinity拼接数据,采用Corset和BUSCO评估质量。基于nr、KOG/COG、Swiss Prot、KO、GO数据库进行注释,E值小于10-5。计算基因表达水平,利用DESeq或TMM标准化readcount数据,使用Benjamini-hochberg方法调整p值。当p<0.05且log2 FC大于1时,认定为差异表达基因,进行GO和KEGG功能富集分析,筛选与附肢发育和腹肢特化相关的基因。
-
① 针对特异SV利用网站CRISPRdirect(
http://crispr.dbcls.jp/ )设计特异性较高的gRNA靶点。②引物合成在擎科生物公司进行。③ PCR反应使用高保真PCR酶(GXL酶),反应体系:10 μL 5×GXL buffer,4 μL dNTP,3 μL上游引物,3 μL下游引物,1 μL GXL酶,29 μL ddH2O。④ DNA纯化回收并检测。⑤ sgRNA的体外转录使用T7体外转录试剂盒。反应体系:10 μL T7 buffer,0.75 μL dNTP,4 μL DNA模板,1 μL T7酶。加好上述试剂并在涡旋仪上混匀后离心,在PCR仪中设置37 ℃、12 h的程序进行反应。⑥去除DNA模板:按照反应产物与DNase比例为20∶1加入,在37 ℃条件下孵育1 h。⑦纯化体外转录产物:按照30∶1配制无水乙醇和醋酸钠混合液1 mL,将回收产物加入混合液中,颠倒混匀后,在-20 ℃条件下沉淀1 h;4 ℃条件下14 500 r/min离心10 min,弃掉上清;用75%酒精洗涤,14 500 r/min离心10 min,弃掉上清;14 000 r/min空离2 min,吸去液体,倒置离心管,以晾干酒精;待酒精挥发干净后,加入20 μL DEPC水放入4 ℃冰箱中过夜溶解,待溶解完全后取1 μL进行浓度测定,标记好浓度后放置-80 ℃冰箱中保存。 -
① 将合成的sgRNA和Cas9蛋白混匀成10 μL的混合液,1 μL sgRNA1,1 μL sgRNA2,1 μL Cas9,用RNase-free H2O补足至10 μL,将混合液放置在PCR仪上,37 ℃孵育15 min。②蚕卵制备:将已经交配6 h的15 ℃催青的野生型Dazao蛾子拆对,将雌蛾放置在蚕卵纸上产卵,把握好产卵时间,在3 h内显微注射完成。③蚕卵消毒:把载玻片放在无水乙醇中浸泡,在火焰上灼烧消毒,之后用浆糊将刚产下的蚕卵粘在载玻片上。④蚕卵显微注射:用拉针器拉毛细玻璃针,将在PCR仪中孵育好的混合液转移到玻璃针中,使用显微注射仪将其注射到蚕卵中。⑤蚕卵封口:取适量的胶水封住注射后蚕卵上的针孔。⑥蚕卵的催青:将注射后的蚕卵置于恒温恒湿培养箱内(温度25 ℃、湿度70%~80%)进行催青,期间维持湿度条件以避免胚胎脱水并定期通风,确保卵壳表面干燥,抑制霉菌滋生。
-
① 用流水清洗配制胶专用玻璃板,并用无水乙醇擦拭玻璃板(防止有杂质存在),清洗好后按照特定顺序组装玻璃板,待组装好后下层用琼脂糖胶封住,30 min后待琼脂糖凝固。② Suryeror丙烯酰胺胶的配制,按配方配制胶浓度为8%的胶块:4.55 mL 5×TBE,2.67 mL Acr Bis(30%),625 μL 80%甘油,150 μL 10% APS(现配),10 μL TEMED,4.55 mL ddH2O。③待配制胶凝固后,对PCR的目的产物进行上样,对有缺失的目的片段对应的PCR产物进行测序,筛选被编辑后该基因功能丧失的蛾子(G0代)进行交配,并对其进行表型观察,继续饲养几代筛选纯合体。
1.1. 家蚕胚胎解剖取材、RNA提取和qRT-PCR
1.2. RNA-seq文库构建及测序
1.3. CRISPR/Cas9介导的基因编辑
1.3.1. sgRNA合成
1.3.2. 显微注射
1.3.3. 编辑效果检测及表型观察
-
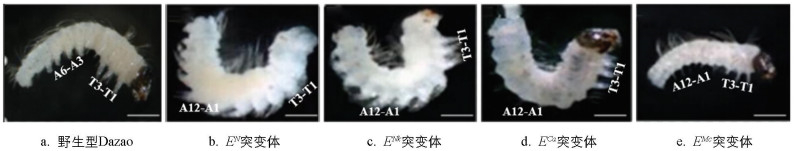
由图 1可知,在野生型Dazao胚胎中,胸部T1-T3体节稳定发育有3对胸足,腹部A3-A6体节则形成4对形态特化的腹足,两类附肢在结构上呈现出显著差异。EN、ENk、ECa和EMc突变体的纯合胚胎均表现出致死表型,且腹部附肢发育异常,其中EN和ENk突变体在腹节异位产生与胸足同源的附肢,而ECa与EMc突变体则完全丧失腹足分化的能力。值得注意的是,所有突变体的胸节均维持正常胸足发育模式,3对胸足的形成未受明显影响。
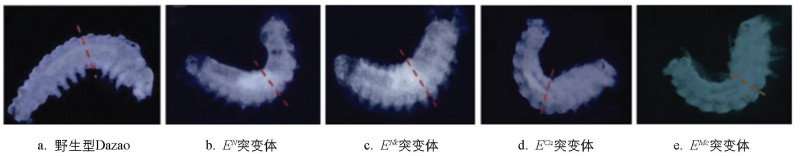
在家蚕胚胎发育的早期阶段,附肢原基已启动形态建成。对野生型Dazao和EN、ENk、ECa、EMc突变体的胚胎进行解剖观察,发现在胚胎催青至腹肢突起出现的这一重要时刻,野生型Dazao与突变体胚胎在腹足的发育上存在明显差异。这一时期被认为是腹足发育的关键阶段,因此该时期的家蚕胚胎可用于筛选附肢的表型修饰基因(图 2)。
-
利用突变体在腹肢突起出现期的胚胎腹部区域进行RNA-seq分析,成功获得了多个转录组数据(表 1)。5个样品获得的原始序列经质控后获得的有效序列,过滤后碱基数量为0.58~0.75 GB,容错率为0.01%,Q20和Q30的值分别在98%和95%以上,GC碱基量均超过41.00%,对Clean reads进行基因组比对分析,其中89.1%~92.9% reads被比对到家蚕参考基因组上,共获得14 622个Unigenes,表明测序结果良好,可用于后续分析。
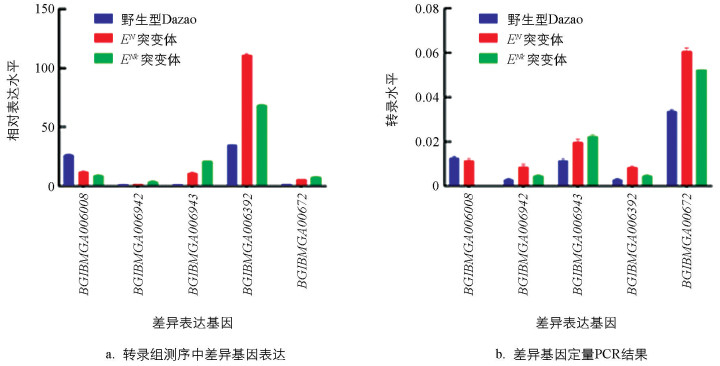
为了分析每个样本的基因表达水平(RPKM,以RPKM表示),我们统计了5个品系不同表达水平的基因数量(表 2)以及单个基因的表达水平(图 3a)。定量分析显示,5个品系中约70%的基因呈现出正常表达水平(RPKM>1),其中约15%的基因达到高表达水平(RPKM>60),揭示这些基因在5个品系间存在保守的高表达特征。
为了验证该转录组测序数据筛选到的差异表达基因,随机选取5个差异表达基因进行荧光定量PCR(qRT-PCR)分析(图 3b),结果表明这些基因的相对表达量与RPKM值变化趋势基本一致,表明该转录组测序结果可靠。
-
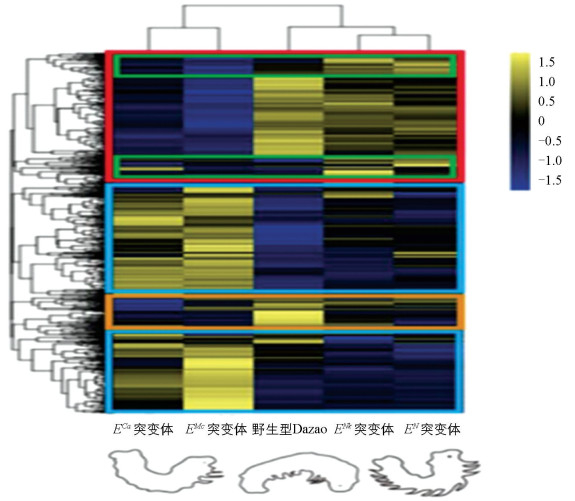
基于各样本的差异基因RPKM值进行层次聚类分析,不同颜色区域代表不同的基因聚类组,显示了同组基因表达模式的相似性,暗示这些基因可能具有相似的功能或参与了相同的生物学过程。根据层次聚类图中差异表达基因的分布模式(图 4)显示,这些基因可被归为4类:第1类基因在长腹足的野生型Dazao和EN、ENk突变体中呈现上调表达,而在不长腹足的ECa和EMc突变体中下调,推测这些基因可能与家蚕腹足的形成与发育相关。第2类基因在野生型Dazao和EN、ENk突变体中下调,而在ECa和EMc突变体中上调,表明它们可能参与了家蚕足发育过程中的负调控机制。这两类基因的表达差异在两个不长足的突变体中尤其明显。第3类基因在EN和ENk中相比于野生型Dazao和ECa、EMc突变体显著上调,提示它们可能在胸足的特化过程中发挥着重要作用。第4类基因在野生型Dazao中的表达量相对于其他品系明显上调,推测其可能是家蚕腹足发育调控的特异性基因。聚类分析不仅揭示了这些基因的表达特征与家蚕足发育的潜在关联,还为进一步研究基因功能及其在家蚕足分化中的具体作用提供了线索。
-
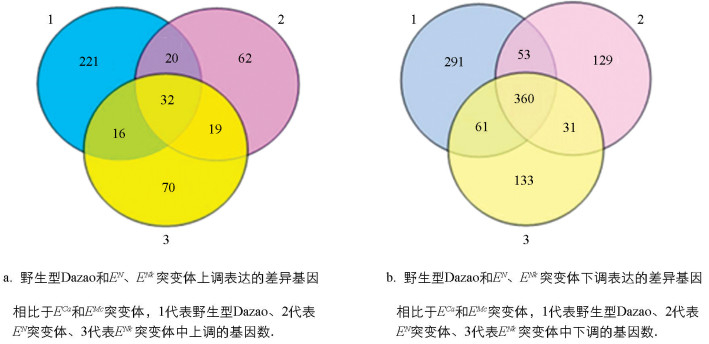
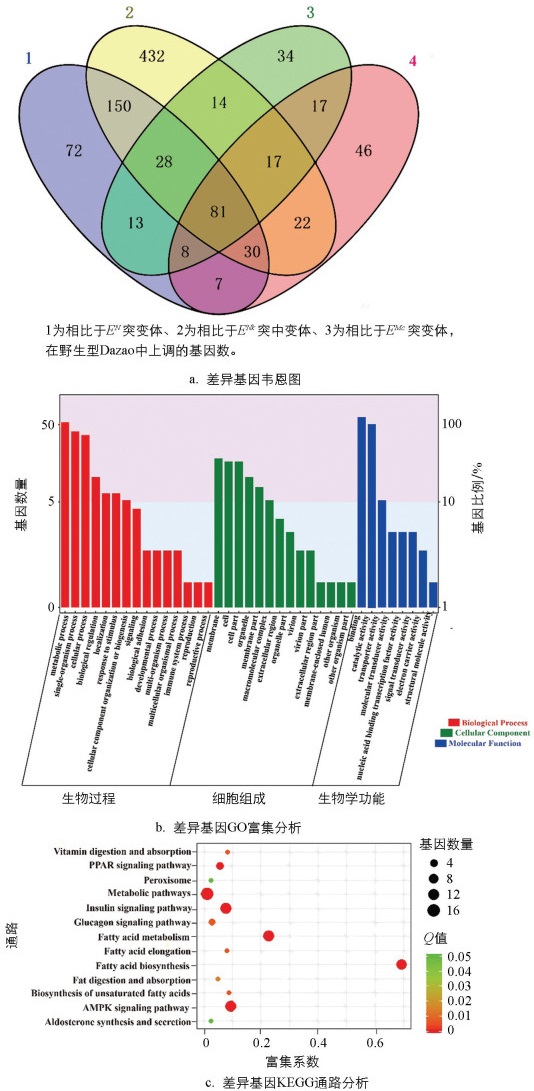
聚类分析结果显示,野生型Dazao和EN、ENk突变体与ECa、EMc突变体的差异基因分别在不同组别聚集。韦恩图分析进一步表明,与ECa和EMc突变体相比,野生型Dazao和EN、ENk突变体中分别有289、133和137个基因呈上调表达,其中有32个基因在这3个品系中共同上调,推测这32个基因可能与家蚕腹足的发育过程相关(图 5a)。此外,与ECa和EMc突变体相比,野生型Dazao和EN、ENk突变体中分别有765、573和585个基因呈下调表达,共有360个基因在3个品系中均下调,推测这些基因可能与家蚕腹部长足的形成和功能相关(图 5b)。综合表明,这些差异基因的表达特征提示它们可能在家蚕特定足部结构的发育过程中发挥着重要作用。
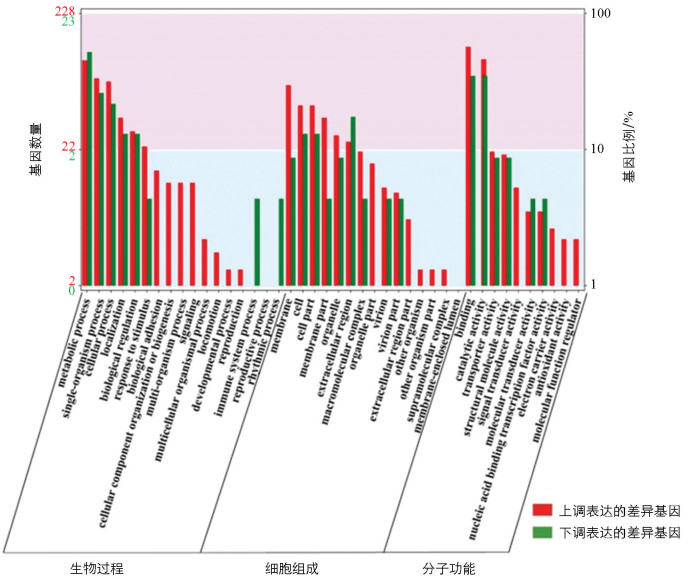
通过对差异表达基因进行GO富集分析(图 6),结果表明,在生物过程类别中,无论是上调基因还是下调基因,都明显富集于代谢过程这一类。值得注意的是,生物黏附(biological adhesion)、细胞成分的组织或生物发生(cellular component organization or biogenesis)、多生物过程(multi-organism process)、信号传导(signaling)、多细胞生物过程(multicellular organismal process)、运动(locomotion)、发育过程(developmental process)、繁殖(reproduction)这些类别仅在上调基因中得到富集,暗示这些生物过程在家蚕足发育的正向调控中可能具有重要的作用。相对而言,免疫系统过程(immune system process)和节律过程(rhythmic process)只在下调基因中富集,表明这些生物过程可能与家蚕足发育的负向调控或其他生物学功能相关。在细胞组成类别中,上调基因主要集中在膜外区域(extracallular region part),下调基因主要富集在膜(membrane part)。在分子功能类别中,可以观察到催化活性(catalytic activity)、蛋白质结合(binding)是上调与下调基因所主要富集的功能类型。这些功能富集结果进一步揭示了家蚕足发育的复杂调控机制,即上调基因富集在多个与发育和细胞组织相关的生物过程中,强调这些基因在家蚕足发育中潜在的关键角色,而下调基因则更多涉及免疫和节律过程,代表不同的生理调节机制。此外,这些差异基因在膜外和膜中的富集分布,提示它们可能参与了细胞间信号传递、细胞外基质重塑及膜受体功能的调控等[21]。
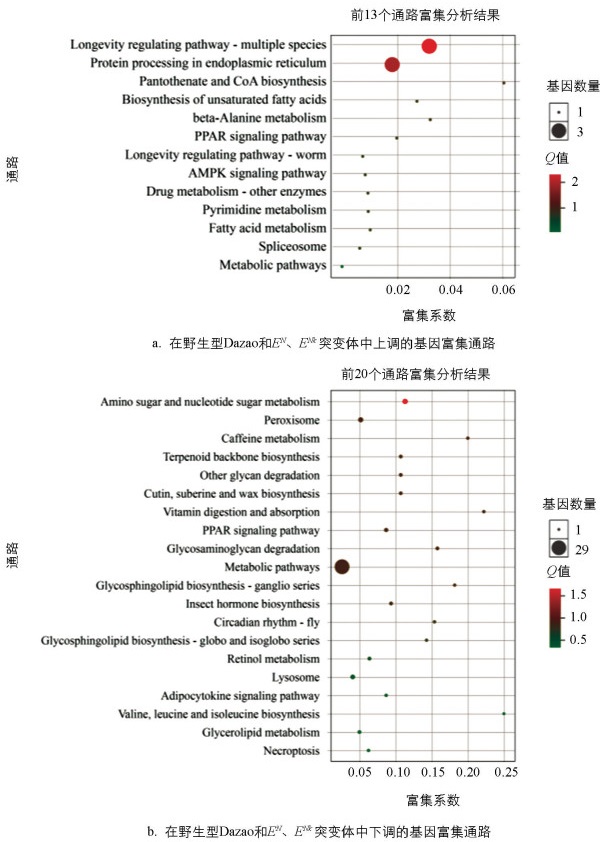
利用KEGG数据库进行差异表达基因的通路富集分析,共得到103条通路,选取富集最显著的33条通路进行分析(图 7)。在生物学过程类别中,我们观察到野生型Dazao和EN、ENk突变体在细胞分裂和细胞周期调控方面有一系列基因呈现出显著上调,暗示这些过程在发育早期阶段可能特别活跃。此外,信号传导和细胞应答等过程也表现出基因表达的显著上调,反映了发育过程中细胞对内外环境变化的敏感性和响应性。在细胞组成方面,细胞核和细胞膜相关基因的表达模式显著变化,表明了细胞在发育过程中可能会重塑其结构以适应不断变化的功能需求。对于分子功能而言,观察到转录调控因子和酶活性相关基因的表达变化,这些变化可能对调控发育过程的基因表达网络至关重要。
结果表明,寿命调节途径、内质网中的蛋白处理、泛酸和辅酶A生物合成、不饱和脂肪酸的生物合成、β-丙氨酸代谢、过氧化物酶体增殖物激活受体(PPAR)信号途径和蛋白激酶AMP活化的蛋白激酶(AMPK)信号途径都显示出显著的富集。这暗示了在特定条件下,这些代谢途径可能起着重要的调控作用,如PPAR信号途径的富集可能与脂质代谢和能量平衡有关,而AMPK信号途径的富集通常与细胞能量应答相关。值得注意的是,代谢途径富集分析常常可以指出疾病状态或生物应答的关键调节点,如咖啡因代谢的富集可能与摄入咖啡因后的生物效应有关,Circadian rhythm-fly途径的富集则可能揭示了生物钟对生物代谢的影响。
将筛选出的差异基因进行功能注释,我们发现在32个上调基因中,有3个热激蛋白、5个酶类、6个激素生长类蛋白、2个疾病相关蛋白,剩下的是未知功能蛋白,其中有3个与附肢发育相关的基因:BGIBMGA012898、BGIBMGA008257、BGIBMGA011483,可作为我们后续研究的目标基因。
-
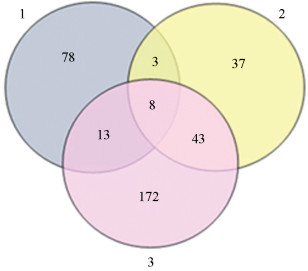
通过分析韦恩图,在EN和ENk突变体中,相较于野生型Dazao和ECa、EMc突变体,分别有102、91和236个基因上调,有8个共同上调的基因(图 8)。对这8个共同上调表达的基因进行功能注释后,发现除基因BGIBMGA001605和BGIBMGA003836被注释为“No hits”外,其余与组织发育、表皮蛋白和转录因子有关,如 Meis1同源盒蛋白基因(BGIBMGA004885)、转录因子Sp9基因(BGIBMGA006943)、水通道蛋白基因(BGIBMGA007757)、TFIID转录起始复合体基因(BGIBMGA009793)、类黏蛋白基因(BGIBMGA011453)等。
视动盲蛋白样(Optomotor-blind protein-like)基因(Omb基因)表达广泛用于果蝇(Drosophila)以及其他模式生物的研究中,Omb基因与成体和胚胎的发育相关,尤其是在神经系统和翅膀的形成中发挥着重要作用。在家蚕的翅膀或腹足等结构发育过程中,该基因的表达调控可能对特定组织的形成和分化显著相关。该基因属于T-box转录因子家族,能够调控下游基因的表达,参与发育、形态形成和细胞分化等生物过程。紊乱型形态发生相关激活因子(Disheveled-associated activator of morphogenesis,DAAM1)是一种与形态发生相关的蛋白质,参与细胞骨架调控、细胞形态和运动等关键发育过程。DAAM1是Wnt/PCP(平面细胞极性)信号通路中的重要成员,特别是在非典型Wnt信号通路中,调节细胞骨架的重塑和细胞迁移。同源框蛋白(Homeobox protein) Meis1是一类包含同源盒的转录因子,在胚胎发育、细胞分化和调控多个基因表达中发挥着重要作用。Meis1基因在人类和其他动物的许多发育过程中均具有关键作用,特别是在造血、神经发育和肢体形成方面。转录因子(Transcription factor) Sp9-like是锌指蛋白家族的一员,通常与细胞分化、器官发育和细胞周期调控等过程相关。Sp9-like蛋白属于Sp家族转录因子的一员,与该家族的其他成员一样,它能够特异性地识别并结合DNA上的特定序列,进而调节与之相关的下游基因的转录活性。在家蚕的肢体形成过程中,Sp9-like转录因子调控肢芽区域细胞的增殖和分化,帮助组织正确形成并维持肢体形态(表 3)。
-
与EN、ENk、ECa和EMc突变体进行比较后,野生型Dazao共筛选出81个共同上调的差异基因(图 9a)。通过基因GO富集分析,这些差异基因被归类为生物过程、细胞组成和分子功能3个类别。结果显示,代谢过程、膜结构及结合相关基因显著富集,提示其可能在家蚕腹足发育中具有关键作用(图 9b)。基于KEGG通路富集分析,筛选到4条显著富集通路:脂肪酸代谢、脂肪酸生物合成、AMPK信号通路和胰岛素信号通路,上述通路均与能量代谢调控密切相关,进一步证实其在腹足发育中的核心作用(图 9c)。
对这些差异基因进行进一步的功能注释,发现基因BGIBMGA006388被注释为Abdominal-A同源蛋白(abdominal A isoform 3 homeobox protein),该基因在家蚕腹部发育中发挥着关键作用,揭示了其在形态形成中的潜在影响。结果表明,代谢相关基因和信号通路在家蚕腹足的发育过程中可能具有重要的调控作用,代谢过程的显著富集提示了代谢活动在足部形成中的重要性,而AMPK和胰岛素信号通路的富集进一步支持了能量代谢在该过程中的关键地位。未来的研究应关注这些差异基因的具体调控机制及其与已知发育通路的交互作用,尤其是BGIBMGA006388的功能验证,以揭示其在家蚕形态发育中的精确角色。
-
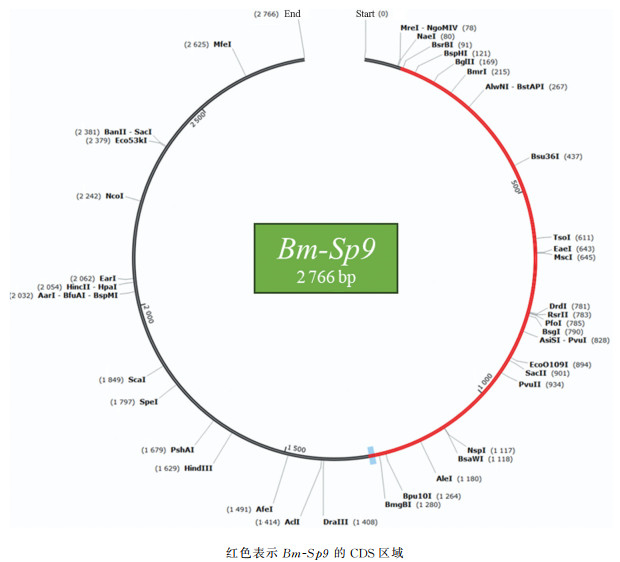
通过同源序列比对分析,Bm-Sp9与果蝇Dm-Sp1的氨基酸序列相似性达61%。从蛋白结构来看,两个蛋白N末端存在由13个氨基酸组成的Sp-box基序,而锌指结构域上游则包含10个氨基酸构成的Btd-box基序(图 10)。尽管这些基序的功能尚未完全明确,但推测Sp-box可能作为抑制因子结合位点,而Btd-box或参与了反激活。值得注意的是,除锌指结构域外,Sp-box与Btd-box在果蝇和家蚕Sp蛋白中均表现出高度序列保守性。基于上述保守特征与序列相似性,我们提出Bm-Sp9与Dm-Sp1可能为功能同源基因。
为了进一步了解Bm-Sp9基因的序列信息,我们从NCBI上调取该基因的mRNA序列进行分析。通过分析发现,该基因的mRNA全长为2 766 bp,其中编码功能域的CDS序列长度为1 218 bp(图 11)。
-
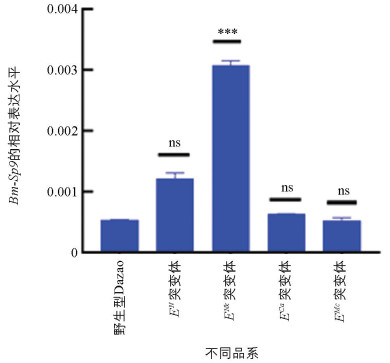
为了进一步探究这些腹肢发育突变体与Bm-Sp9基因表达之间的关系,我们通过荧光定量PCR技术检测了Bm-Sp9基因在野生型Dazao和EN、ENk、ECa、EMc突变体胚胎发育第5 d时的表达量。结果显示,在胚胎附肢发育的关键时期,Bm-Sp9在ENk突变体中表现出显著的上调(图 12)。这表明Bm-Sp9基因的表达可能在家蚕胸足的发育过程中起着重要作用,我们推测其参与了胸足形态的形成或功能的调节。
-
为了进一步确认该基因对突变体ENk胸足发育的影响,利用CRISPR/Cas9介导的基因编辑技术在ENk突变体中对Bm-Sp9基因进行敲除。
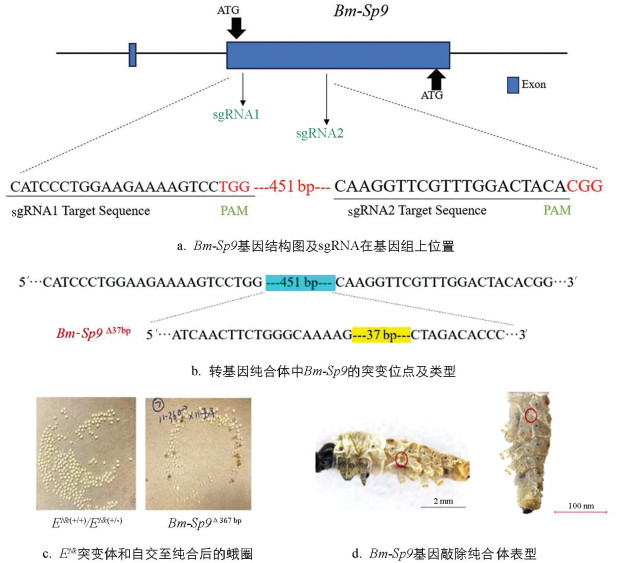
首先,利用突变体ENk的基因组序列设计2个敲除靶位点sgRNA1、sgRNA2(图 13a);随后,将体外转录的2个sgRNA产物与Cas9蛋白均匀混合后,37 ℃孵育获得Cas9 RNP,注射于刚产下的ENk突变体蚕卵中。由于突变体ENk纯合致死现象,在注射的蚕卵中一共孵化了64头,G0代幼虫未发现附肢异常的个体,饲养至出蛾,剪取蛾子的部分翅膀进行基因组提取,并根据sgRNA位点设计出合适的检测引物检测基因的编辑效果。结果显示:存在1种不同形式的序列突变类型,为缺失37 bp(Bm-Sp9 Δ37 bp),将其自交至纯合,观察其表型变化(图 13b)。纯合体表型回复为T1-T3有3对胸足、A4-A6有3对腹足,与野生型Dazao表型一致。图 13c为ENk突变体和自交至纯合后的蛾圈。值得注意的是,纯合体Bm-Sp9 Δ37 bp的A3环节处缺失1只腹足(图 13d)。
2.1. 家蚕野生型Dazao及EN、ENk、ECa、EMc突变体表型观察及取材
2.2. 转录组数据统计与评估
2.3. 差异表达基因分析
2.3.1. 家蚕足发育相关基因的筛查
2.3.2. 家蚕胸足特化相关基因筛查
2.3.3. 家蚕腹足发育相关基因筛查
2.4. 差异表达基因的功能验证
2.4.1. Bm-Sp9的生物信息学分析
2.4.2. Bm-Sp9基因表达模式分析
2.4.3. Bm-Sp9基因的功能验证
-
昆虫作为自然界中进化最为成功的生物类群之一,其适应性优势与附肢的多样化演变密不可分。尤为关键的是,胸部与腹部附肢通过形态和功能的特化,形成了显著的空间分工,如在完全变态昆虫的幼虫阶段,不仅胸足负责运动与抓握,其腹部还发育出特化的腹足结构。这种形态与功能的协同分化(如胸足呈现节段性关节,而腹足多具吸盘或钩状末端)使昆虫能够针对取食、攀附、感知等不同需求进行精准适应。本研究利用高通量转录组测序技术[22-24],揭示了Hox基因在家蚕腹足表型修饰中的下游调控网络。研究选取了家蚕品种野生型Dazao和EN、ENk、ECa、EMc突变体,在腹足突起发育阶段的胚胎腹部组织作为样本,成功获取了126.70 Gb的高质量序列数据。基于获得的转录组数据,通过差异基因的聚类分析,将差异表达的基因分为4个主要类别:第1类基因在野生型Dazao和EN、ENk突变体中表现为上调(相较于ECa和EMc突变体)。第2类基因在野生型Dazao和EN、ENk突变体中表现为下调(相较于ECa和EMc突变体),这两类基因的表达模式可能与家蚕足的形成和发育过程紧密相关。第3类基因仅在EN和ENk突变体中表达上调,推测与胸足特化功能相关。第4类基因在野生型Dazao中表达量高于其他4个品种,可能参与了腹足的发育过程。这一分析方法有效地展示了不同基因类别之间的交集和独特性,为后续的功能验证和分子机制研究提供了重要线索。
通过GO富集、KEGG通路分析及基因功能注释,初步筛选了可能参与调控家蚕腹足表型的关键基因(表 3)。特别注意到,Omb基因在野生型Dazao和EN、ENk突变体中表现出显著上调,其在果蝇中作为Dpp基因的下游因子在肢体发育中起作用,推测家蚕的同源基因BGIBMGA012898可能参与了腿部形态的形成[25]。Hox基因家族,作为编码含有同源结构域转录因子的一组基因,在动物胚胎的体轴建立中发挥着核心作用[26-27],与之相辅的Meis1基因,属于TALE超家族,是已知的多种发育过程中的关键转录因子,在鸡和小鼠的肢体近端至远端轴(P-D)的发育中扮演着重要角色[28]。Meis1的异位表达可引起远端肢体结构向近端结构转化。在家蚕的附肢发育中,Meis基因通过与其他基因相互作用调控附肢特异的细胞分化和发育,如Meis基因与Pbx基因协同作用,共同参与调控下游目标基因的表达。在肢体生长过程中不可或缺的Sp转录因子家族,在进化上表现出功能的保守性。研究证实,果蝇Sp家族成员btd与Sp1编码锌指转录因子,通过激活hdc及Dll基因,驱动腹侧成虫盘的形成与生长调控[29]。RNA干扰实验显示,当btd与Sp1基因功能失活时,果蝇腿与触角尺寸显著缩小[30],表明二者对附肢发育具有剂量依赖性调控作用。值得注意的是,btd在背侧成虫盘(如眼、翅及平衡棒)中的异位表达可诱导腹侧结构(触角、腿)的异常生成,提示其具有组织特异性形态建成的指导能力。进一步比较分析发现,家蚕同源基因BGIBMGA004885与BGIBMGA006943在EN、ENk突变体中呈现出显著上调的表达特征,暗示其可能通过保守的调控机制参与了附肢的发育过程。Sp家族基因通过影响细胞的增殖和分化过程参与调控附肢的形态建成。此外,Sp9可能与Hox基因以及其他信号传导通路交互,共同决定附肢的最终形态,推测它们在家蚕胸足发育和特化中起到了重要作用。研究显示,Hox基因家族成员abd-A在节肢动物腹部附肢发育中具有调控多样性[31],在果蝇中,该基因通过下调Dll基因的转录活性,最终导致幼虫阶段腹足结构的发育停滞[32-33]。Dll基因是调控附肢发育的另一个重要基因,它在许多动物的附肢形成中发挥着核心作用。在家蚕中,Dll表达在发育早期附肢的远端部分,对于确定附肢的远端结构特别是足部的形成至关重要,而abd-A则促进了家蚕腹足的发育[33]。abd-A在野生型Dazao中相比于ECa、EMc、EN、ENk突变体上调表达,证明了其在正常腹足发育中的功能。dac基因在昆虫的附肢发育中也非常关键,它在家蚕中主要表达于附肢的中部,与附肢的长度和身体比例调控相关。dac与Dll、hox基因之间存在互作,共同细化附肢的形态和功能。exd和hth两个基因编码的蛋白质在昆虫中与Hox蛋白互作,帮助执行Hox基因的发育指令。在家蚕中,exd和hth的表达模式影响了附肢的基部至远端的形态发展。通过转录组分析鉴定出的这些附肢(胸足和腹足)发育相关基因仍需要深入研究,以确定其具体功能和调控方式。
家蚕的附肢发育调控网络包括多个信号途径,如Wnt、Hedgehog (Hh)、Decapentaplegic (Dpp)和Epidermal growth factor (EGF)信号途径。这些信号途径通过激活或抑制特定的转录因子和基因,协同工作以精确控制附肢各部分的形态和大小。例如,Wnt信号途径在促进附肢远端结构的形成中尤为重要,而Hh和Dpp信号途径则在整个附肢的轴向增长和分化中发挥作用。本研究对野生型Dazao和ECa、EMc、EN、ENk突变体胚胎进行了转录组分析,获得了一大批差异表达基因,可为深入阐明这些基因的功能、了解鳞翅目昆虫附肢多样性打下基础。