


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
茶树(Camellia sinensis)属山茶科山茶属的常绿木本植物,是我国重要的经济作物[1],其生长状况直接影响到茶叶的产量和品质。茶树根系的健康是保证其生长的基础,根系内生微生物群落,尤其是内生细菌和真菌,对根系健康生长及水分、养分的吸收利用均具有重要影响。
植物内生菌是指能够在宿主植物细胞内(或细胞间)度过整个(或部分)生命周期而不引起植物病理性病变的一类微生物[2]。常见的植物内生菌类群主要包括内生真菌、内生细菌和内生放线菌[3]。研究表明,内生细菌能够通过产生植物生长调节物质如吲哚乙酸(IAA)等,促进茶树根系的生长,增强其对营养的吸收能力[4]。此外,内生真菌也能够影响茶树的代谢途径,从而影响茶叶的化学成分和品质[5]。在茶树生长过程中,茶树能够与其内生菌相互作用,提高茶树对病原微生物的抗性。黄晓琴等[6]从茶树健康组织上筛选鉴定到解淀粉芽孢杆菌Y1,能够显著抑制冰核细菌生长的内生菌,防治茶树霜冻害。张林平等[7]从健康油茶叶片中分离鉴定得到6株内生细菌菌株,其对油茶炭疽病抑制率最高可达到82.4%。宋维兴[8]研究证实,在Cd-LDPE(镉-低密度聚乙烯)复合污染下,接种植物内生菌增加花生地下部生物量,显著提高光合作用和SPAD值,显著降低了细菌群落多样性和丰富度。Yan等[9]发现在防草膜覆盖下,根系土传病原菌尖孢镰刀菌的相对丰度显著降低。
本试验以云南省澜沧县阿木戛村茶园中不同除草方式下的茶树根系内生菌分布及拮抗活性为研究内容,将分离纯化获得的内生菌株,通过与植物病原真菌拮抗活性筛选及分子生物学鉴定,探究其潜在的生物防治价值,旨在获得具有优良抑菌活性菌株,为茶叶新型活性抑菌物质和新型生物农药的研发提供理论依据。
全文HTML
-
采样时间为2022年4月,采样茶园位于云南省澜沧县阿木戛村,3种处理方式茶园位于同一区域。海拔为900~1 640 m,年平均气温为16.5 ℃,年降雨量为1 673 mm,为茶树的生长提供了良好的气候条件。在采样区域,选择了3个具有代表性的茶园样地为研究对象,栽种的茶树品种均为地方群体种(景迈群体种),3个取样茶园采用了不同的杂草管理方式,A样点使用防草膜覆盖除草的方式,防草膜在2019年12月前完成铺设,覆盖时间在两年以上,防草膜的材质为聚乙烯;B样点采用人工耕作除草的方式;C样点则是不除草的管理。除不同除草方式外,所有样点茶园都采取了相同的栽培措施,如种植遮荫树、不施用化肥和不打农药。3个取样茶园分布地理信息及除草方式见表 1。在每个位点采用对角线法选取3株健康茶树进行根系组织的采集,总共获得9个根系样本。将采集的样本放置于冰上,24 h内带回实验室进行处理。
-
PDA培养基(用于菌株筛选):马铃薯(去皮)200 g,蔗糖(或葡萄糖)20 g,蒸馏水1 000 mL。配制方法如下:将马铃薯去皮,切成约2 cm2的小块,放入1 500 mL的烧杯中煮沸30 min,注意用玻棒搅拌以防糊底,然后用双层纱布过滤,取其滤液加糖,再补足至1 000 mL;Martin培养基(用于真菌分离):K2HPO4 1 g,MgSO4·7H2O 0.5 g,蛋白胨5 g,葡萄糖10 g,1/3 000孟加拉红水溶液100 mL,蒸馏水900 mL,121 ℃湿热灭菌30 mins,待培养基融化后冷却至55~60 ℃时加入链霉素(链霉素含量为30 μg/mL);牛肉膏蛋白胨培养基(用于细菌分离):牛肉膏3 g,蛋白胨10 g,NaCl 5 g,蒸馏水1 000 mL,pH值7.4~7.6。
-
选择5株植物病原真菌作为对峙试验的供试病原菌:卷枝毛霉菌(Mucor circinelloides)(CGMCC 3.215)、茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)、茶树炭疽病(Colletotrichum camelliae)、尖孢镰刀菌(Fusarium oxysporum)(CGMCC 3.363 8)、白地霉(Geotrichum candidum)菌株,均由云南农业大学及云南农业科学院茶叶科学研究所提供,并于中国普通微生物菌种保藏管理中心(
https://cgmcc.net )保存。 -
采集的茶树根系样本经去除表面土壤与杂质等预处理后,依次进行表面消毒处理:用无菌水冲洗30 s,浸入75%乙醇1 min,转入2.5% NaClO溶液浸泡3 min,再次用75%乙醇浸泡1 min,最后用无菌水冲洗3次。取根系中段约1 cm部位,纵切后使切口最大限度接触培养基,分别接种于Martin培养基和牛肉膏蛋白胨培养基。样本置于28 ℃黑暗环境培养3~7 d,记录菌落形态、生长速度和颜色差异。
从初筛菌落中挑取边缘生长良好且未污染的部分,转移至培养基进行纯化,筛选出具备内生真菌与细菌特征的菌株并编号,为后续试验提供明确样本。向试管加入约8 mL PDA培养基,灭菌后倾斜冷却成斜面。使用无菌接种针从纯化平板转移菌落菌丝接种至培养基斜面,培养完成后,试管置于4 ℃冷藏保存,延缓菌株代谢以长期保藏资源。
-
采用五点对峙培养法测定了内生菌对病原菌的拮抗作用[10]。选择5种植物病原菌作为对峙试验的供试病原菌,在直径为9 cm的PDA平板正中央接入直径为5 mm的供试病原菌菌饼,同时分别在平板距供试菌菌饼2.5 cm处接入4个相同大小的内生菌菌饼,以只接供试病原菌、不接内生菌的平板作为对照,每个处理重复3次,25 ℃条件下培养。每24 h观察1次,从对峙培养第3 d起测量病原菌菌落直径和抑菌带距离,统计抑菌率。
抑菌区距离(D)为内生菌菌落与被试病原菌菌落之间的无菌区宽度。
-
将筛选所得拮抗活性较好的菌株进行纯化培养,再进一步进行形态观察,记录其菌落特征、细胞形态等信息之后再进行分子生物学鉴定。使用试剂盒提取拮抗活性较好的菌株基因组DNA,采用通用引物36F(5′-TCTGGCTCAGGATGAACGC-3′)和1492R(5′- GGTTACCTTGTTACTT -3′)扩增活性细菌菌株的16S rRNA基因序列,使用通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)扩增活性真菌菌株的内部转录间隔区(ITS),对具有显著抑制活性的细菌或真菌进行分子生物学鉴定。对所得PCR产物进行测序,并使用National Center for Biotechnology Information (NCBI)数据库(
http://www.ncbi.nlm.nih.gov )进行BLAST (Basic Local Alignment Search Tool)检索,与GenBank中存储的其他序列进行比对。这些序列在相同的方向,即5′~3′,保存为FASTA格式,并上传到MEGA X软件(Molecular Evolutionary Genetics Analysis X)上,将序列对齐并修剪到合适的长度以获得保守区域。然后将这些修剪过的序列转换为MEGA X格式并保存以用于Neighbor-joining系统发育树的构建[11]。
1.1. 试验材料
1.1.1. 茶园除草方式与茶树根系采样
1.1.2. 培养基
1.1.3. 供试植物病原真菌
1.2. 试验方法
1.2.1. 茶树根系内生菌分离与纯化培养
1.2.2. 拮抗活性菌株筛选
1.2.3. 活性菌株分子生物学鉴定及系统发育树构建
-
从不同除草方式下茶树的健康无症状根系组织中分离得到37株内生菌(表 2)。其中,分离自防草膜覆盖除草、人工耕作除草和不除草处理的分别有14株、13株和10株,分离自Martin培养基和牛肉膏蛋白胨培养基的分别有24株和13株。
-
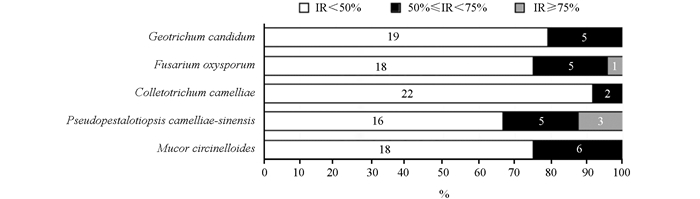
从3种除草方式茶园茶树根内分离出24株内生真菌,为了评估其与植物病原真菌的拮抗能力,24株内生真菌与5种植物病原真菌进行了对峙培养,其抑菌率见表 3。根据拮抗作用强弱分为三级:强(抑制率≥75%)、较强(50%≤抑制率<75%)和弱(抑制率<50%)[12]。由表 3可知,11株内生真菌对供试病原真菌具有较强的拮抗作用,占内生真菌的45.83%。其中,对至少1种供试病原真菌的抑制率达75%以上有4株(菌株A1-8、C1-2、C1-3和C1-5),具有强拮抗活性。
图 1表明,24株内生真菌对卷枝毛霉(Mucor circinelloides)抑制率为50%~75%的内生真菌有6株,占25%;对茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)抑制率为50%~75%的内生真菌有5株,占20.8%,抑制率75%以上的内生真菌有3株,占12.5%,其中,C1-2表现出的抑制率最高,达到84.52%;对茶树炭疽病(Colletotrichum camelliae)抑制率为50%~75%的内生真菌有2株,占12.5%;对尖孢镰刀菌(Fusarium oxysporum)抑制率为50%~75%的内生真菌有5株,占20.8%,抑制率在75%以上的内生真菌共1株,占4.1%,达到76.92%(A1-8);对白地霉(Geotrichum candidum)抑制率为50%~75%的内生真菌共5株,占20.8%。
-
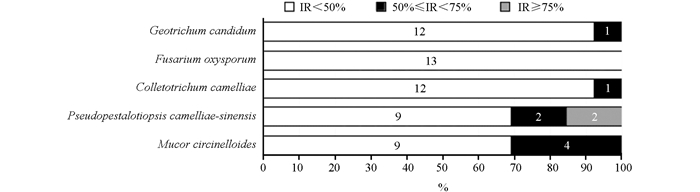
从3种除草方式茶园茶树根内纯化分离获得13株内生细菌(表 4)。由表 4可知,6株内生细菌对供试病原真菌具有较强拮抗作用,占46.15%。菌株A2-2和B2-2对茶树轮斑病菌的抑制率超过了80%,显示出强抑制作用。图 2表明,13株内生细菌对尖孢镰刀菌(Fusarium oxysporum)显示出弱抑制效果,其抑制率均低于50%;对卷枝毛霉(Mucor circinelloides)抑制率为50%~75%的内生细菌共4株,占30.77%;对茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)抑制率为50%~75%的内生细菌共4株,占30.77%,抑制率在75%以上的内生细菌共2株,其中,B2-2抑制率最高,达到86.76%;对茶树炭疽病(Colletotrichum camelliae)抑制率为50%~75%的内生细菌共1株,占15.38%;对白地霉(Geotrichum candidum)抑制率为50%~75%的内生细菌共1株,占7.70%。结果表明,不同内生细菌对不同病原真菌的拮抗作用和强度均有所不同。
-
根据各个菌株对峙培养及拮抗能力试验结果,共有11株内生真菌和5株内生细菌菌株对至少1种病原菌的抑制率在50%以上,属于拮抗活性菌株。其中,部分内生真菌和细菌菌株与病原菌株的拮抗效果如图 3所示。
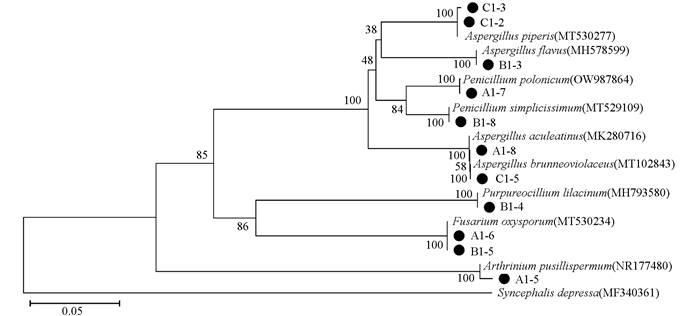
通过菌株测序及分子生物学鉴定,结合GenBank序列比对结果,对11株内生真菌进行分类地位鉴定,并基于其rDNA-ITS序列进行了系统发育分析如图 4,其中,菌株Syncephalis depressa(GenBank登录号MF340361)为外源种。11株活性菌株分别隶属于9个真菌种类,即青霉菌属2个种:Penicillium polonicum(A1-7)和Penicillium simplicissimum(B1-8);曲霉菌属有4个种:Aspergillus piperis(C1-2,C1-3)、Aspergillus flavus(B1-3)、Aspergillus aculeatinus(A1-8)、Aspergillus brunneoviolaceus(C1-5);镰刀菌属1个种:Fusarium oxysporum(A1-6,B1-5);紫孢霉属1种:Purpureocillium lilacinum(B1-4);节菱孢属1种:Arthrinium pusillispermum(A1-5)。
从防草膜覆盖茶树根内分离出的曲霉菌属A1-8对4株病原真菌的抑制率达到50%以上,其中,对Fusarium oxysporum抑制率达到76.92%,属于强拮抗作用;从人工耕作除草茶树根内分离出的曲霉菌属B1-3对4株病原真菌的抑制率达到50%以上;从对照不除草茶树根内分离出的曲霉菌属C1-5对5株病原真菌的抑制率达到50%以上,其中,对Pseudopestalotiopsis cameliae-sinensis抑制率达到80.40%,属于强拮抗作用。同时,曲霉菌属C1-2和C1-3对Pseudopestalotiopsis cameliae-sinensis的抑制率分别达到84.51%和80.69%。
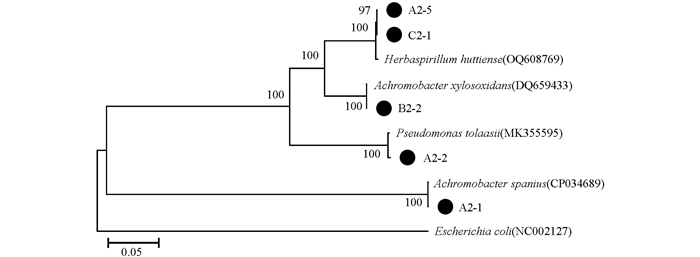
5株内生细菌经过形态学特征和GenBank序列比对分析,其分子生物学鉴定及菌株的16S rRNA序列系统发育树构建分析如图 5,其中,菌株Escherichia coli(NC002127)为外源种。5株活性菌株分别隶属于4个菌种,即假单胞菌属1个种:Pseudomonas tolaasii(A2-2);草螺菌属1个种:Herbaspirillum huttiense(A2-5,C2-1);无色杆菌属2个种:Achromobacter xylosoxidans(B2-2)、Achromobacter spanius(A2-1)。从防草膜覆盖组中分离出的无色杆菌Achromobacter spanius(A2-1)和假单胞菌Pseudomonas tolaasii(A2-2)对3株病原真菌的抑制率达到50%以上,其中,A2-2对Pseudopestalotiopsis cameliae-sinensis抑制率达到80.39%,属于强拮抗作用;从人工耕作除草处理中分离出的无色杆菌属Achromobacter xylosoxidans(B2-2)对Pseudopestalotiopsis cameliae-sinensis抑制率达到86.76%。
2.1. 不同除草方式下茶树根系内生真菌与内生细菌的分离
2.2. 茶树根系分离的内生真菌与病原菌拮抗活性的测定
2.3. 茶树根系分离的内生细菌与病原菌拮抗活性的测定
2.4. 拮抗活性菌株的鉴定
-
茶树各种病虫害滋生与流行正威胁着世界各地茶叶的可持续发展,而内生菌与宿主植物之间能够通过建立密切的互惠共生关系[13],可诱导植物抗病性从而达到防治植物病害的目的。同时,防草膜可以通过相应的覆盖效果抑制茶树病害。首先,其物理屏障作用阻止可能携带不同疾病孢子的雨水或灌溉水进入土壤[14];其次,防草膜覆盖可以改变茶园土壤及茶树根际的温湿度与养分状况等条件,进而影响内生菌在茶树根内定殖与分布。在拮抗菌的分离鉴定中,防草膜处理下鉴定出假单胞菌(Pseudomonas tolaasii),对茶树常见病原菌茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)的抑制率达80.39%。假单胞菌是一种广泛用于防治植物土传病害的生物防治剂[15],防草膜可能对竞争生态位的有益生物提供定殖位来抑制病原体[16],从而达到防治植物病害的目的。
镰刀菌是一种普遍存在的土传真菌[17],其在生长和代谢过程中也会产生一些新兴的真菌毒素,这可能导致植物—土壤负反馈[18]。不仅如此,真菌毒素会污染作物,长期接触会对茶树健康造成危害[19]。本研究也证实在拮抗真菌的分离鉴定中,防草膜处理中分离出对常见土传病害尖孢镰刀菌(Fusarium oxysporum)拮抗效果较好的青霉菌(Penicillium polonicum,70.19%)和曲霉菌(Aspergillus aculeatinus,76.92%),表明防草膜处理茶树根内存在相应的内生菌可以减少病原菌的积累和土壤病害的发生,增加茶树根系内生菌株多样性和抗菌能力。
在农业可持续发展进程中,微生物降解农药残留(特别是敌草隆),是修复环境污染的有效方法之一[20]。无色杆菌(Achromobacter xylosoxidans)作为一种新型生物防治材料,对多种植物病原真菌具有广谱抗性,对小麦赤霉病有较好的防治效果[21],同时能够高效降解敌草隆[22]。本研究从人工耕作除草茶树根系中分离出的无色杆菌(Achromobacter xylosoxidans,B2-2)和淡紫紫孢菌(Purpureocillium lilacinum,B1-4)对茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)的抑制率达86.76%和60.29%。淡紫紫孢菌(Purpureocillium lilacinum)对黑色聚乙烯地膜有较好的降解能力[23]。虽然防草膜覆盖会导致土壤中存在大量的塑料碎片及污染问题,但是,人工耕作除草茶树根中也分离出淡紫紫孢菌(B1-4),说明茶树根内普遍分布有对地膜具备降解能力的淡紫紫孢菌(Purpureocillium lilacinum),它有可能成为未来茶园土壤污染防治的研究方向。
本研究从不除草茶树根系中鉴定出两种曲霉菌(Aspergillus piperis,C1-2;Aspergillus brunneoviolaceus,C1-5)对茶树轮斑病(Pseudopestalotiopsis camelliae-sinensis)的抑制率均在80%以上。这两种内生曲霉菌的发现,表明茶树根系内生菌株能够有效抑制病原真菌的生长,有助于茶园病害的绿色防控。未来的研究将进一步改进茶树根系内生菌株的分离技术、优化培养基和培养条件,拓宽菌种的分离范围,深入探讨这些内生菌株的生物防治潜力,为茶叶领域的新型活性抑菌物质和生物农药的研发提供菌剂资源。
-
本研究从3种不同除草处理的茶树根系中分离出具有拮抗活性的内生菌株。防草膜覆盖处理分离出4株内生真菌和3株内生细菌,其中Aspergillus piperis和Aspergillus brunneoviolaceus对茶树轮斑病的抑制率达80%以上,表现出强拮抗作用;人工耕作除草处理分离出的Achromobacter xylosoxidans和Purpureocillium lilacinum分别对茶树轮斑病的抑制率为86.76%和60.29%。防草膜处理对多种病害,尤其是土传性病害尖孢镰刀菌表现出较好防治效果。研究结果为茶园病害防控及微生物农药研发提供了科学依据,有助于推动茶园可持续发展。