


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
水稻作为世界3大粮食作物之一,对于维持国家粮食安全和世界和平具有重要意义。水稻最主要的作用便是利用其丰富且易消化的淀粉作为主食,为人类提供日常的能量所需。在世界范围内,大米提供了27%的膳食能量供应和20%的膳食蛋白质摄入。仅在亚洲,就有超过20亿人从大米及其衍生产品中获得60%~70%的能量摄入。据联合国粮食农业组织(
https://www.fao.org/faostat/zh/#data )2023年数据显示,中国作为世界第一人口大国,水稻总产为世界第11,因此对于养活14.1亿人口和保障粮食战略安全的中国,提高产量仍然是水稻育种的重点。水稻产量的关键决定因素包括植物形态、穗结构和粒型特征等。这些产量特性相辅相成,共同决定总产量。就单株水稻而言,分析水稻植株形态(如分蘖数、株高、分蘖角度、叶形、茎叶角度和穗类型)和籽粒形成(如花/穗发育和种子发育),对水稻产量性状的分子设计和育种都有重要意义。这项工作需要对控制水稻主要产量性状的相关基因的复杂调控网络进行全面分析,挖掘关键产量相关基因或数量性状位点(QTL)。分子标记辅助选择在水稻育种中的应用已有多年,促使了染色体片段代换系(chromosome segment substitution lines,CSSL)的构建和许多QTL的鉴定[1]。迄今为止,许多与籽粒长度相关的QTL已在水稻中克隆,包括GS3、qGL3、GL4、TGW6、GL7等。GS3是水稻中第一个调节粒长和粒质量的QTL[2-3]。qGL3/GL3.1编码PPKL蛋白磷酸酶家族的丝氨酸/苏氨酸磷酸酶,可通过去磷酸化底物调节细胞周期蛋白,从而控制水稻籽粒的长度和产量[4]。GL4位于4号染色体上,来自长粒的非洲水稻(Oryza glaberrima Steud.),能够调节外层和内层胶质的纵向细胞伸长[5]。TGW6编码具有吲哚-3-乙酸(IAA)-葡萄糖水解酶活性的新型蛋白质,日本晴tgw6等位基因通过控制籽粒IAA供应和限制细胞数量调控水稻的籽粒大小[6]。GL7是控制水稻粒长的主要QTL,位于7号染色体上,编码与拟南芥LONGFOLIA同源的蛋白质。上调GL7的表达水平,使颖壳细胞纵向伸长,从而增加水稻粒长并改变其外观品质[7]。GS5编码丝氨酸羧肽酶,正向调节籽粒大小[8],GS5启动子区域中的两个关键SNPs影响其在幼穗中的表达差异,进一步决定了籽粒大小的差异[9]。GLW7编码OsSPL13蛋白,主要通过增加细胞体积使籽粒变大[10]。GS9编码未知保守功能域的蛋白质,通过改变细胞分裂来调节籽粒形状[11]。qLGY3编码转录因子OsMADS1,是G蛋白β-γ二聚体下游的关键效应子能够产生更长的籽粒[12]。OsLG3是ERF家族转录因子的成员,能正向调控水稻籽粒长度,对籽粒品质无影响[13]。GL12可能参与了调节粒长和粒质量的GS2通路,通过提高灌浆率和受精后颖壳的细胞长度来提高粒长与粒质量[14]。综上所述,通过QTL或突变基因的克隆对水稻粒型等重要性状的遗传机制加以解析,对于水稻的育种和栽培具有极其重要的意义。
然而,这些机制间还存在诸多知识间隙,许多假设也还需进一步证明,许多微效QTL还没有鉴定出来,而微效QTL的鉴定需要更精细的次级作图群体才可完成[15]。这些次级作图群体的构建对后基因组时代的设计育种和微效基因的遗传解析是非常必要的。染色体片段代换系(CSSL)就属于这样的群体。CSSL通常以生产上优良纯合品种作为受体亲本,通过高代回交结合全基因组分子标记辅助选择(MAS)获得,每个CSSL携带来自供体的1个或少量来自供体亲本的代换片段[16]。当CSSL仅含唯一1个供体亲本的代换片段时被称为单片段代换系(single segment substitution lines,SSSL)。利用SSSL进行QTL定位具有明显的优势,其准确度和灵敏度较高,定位结果实用性强,可直接用于育种实践[17],但单片段代换系的构建需多年连续不断的选择才能完成。本研究以含4个代换片段CSSL-Z454的衍生系进行相关性状的QTL定位,构建单片段代换系(SSSL)与双片段代换系(DSSL)进行QTL验证和QTL间的上位性互作分析,旨在为后续新QTL的图位克隆及分子机制解析奠定基础,同时也为水稻分子设计育种提供可靠的遗传信息和遗传资源。
全文HTML
-
主要试验材料为含4个代换片段的CSSL-Z454。Z454是通过分子标记辅助选择法从日本晴与6代换片段CSSL-Z744杂交后代F4代中选育而成的。Z744是以日本晴为受体亲本、自育籼稻恢复系西恢18为供体亲本,通过全基因组429个分子标记辅助选择技术结合高代回交和自交选育而成[18]。QTL定位群体是由日本晴与Z454杂交构建的由130个单株组成的次级F2代分离群体,次级片段代换系构建材料为依据QTL定位结果从F2代分离群体中选出的9个单株。
-
2021年6月,在西南大学水稻研究所实验基地利用受体亲本日本晴与Z454杂交获得杂交种。同年8月种植于海南陵水基地,收获日本晴/Z454的F1代种子。2022年3月,于重庆实验基地播种日本晴、Z454与日本晴/Z454的F1代种子(自交产生F2代)。同年4月中旬以株距16.5 cm、行距26.4 cm为标准,移栽亲本30株、F2代130株至同一试验田中。2023年3—4月中旬,于重庆实验基地种植日本晴、Z454和从F2代中选出的9个单株(种植为9个株系)各30株,种植方式同2022年,参照重庆当地种植方法管理。
-
待7月中旬籽粒完全成熟后,选取日本晴、Z454小区内中间2行3~7株共10个单株,及F2代130株进行收割。参照Wang等[19]的方法对亲本与F2代群体的株高、有效穗数、一次枝梗数、二次枝梗数、穗长、粒长、粒宽、长宽比和千粒质量共9个性状进行考察,使用Excel 2016进行平均数、标准差计算,对日本晴和Z454各性状等进行t检验。
-
移栽2周后,从每个F2代单株及日本晴、Z454和西恢18中取2片叶,通过CTAB法提取DNA作为模板,以Z454代换片段中的10个多态性SSR标记作为PCR的引物,依据Zhao等[20]描述的方法进行PCR扩增与10%聚丙烯凝胶电泳。将受体亲本日本晴的带型读作“-1”,Z454的带型读作“1”,杂合带型读作“0”;然后对F2代130株的基因型,结合对应的表型数据,利用SAS 9.3统计软件的混合线性模型(mixed linearmodel,MLM)法进行QTL定位,以阈值p<0.05判断是否存在控制某一性状的QTL[20]。
-
根据2022年的QTL定位结果,从F2代中选出含有目标QTL的代换标记和0~1个杂合标记的9个单株,在2023年种植为9个株系自交,每一个株系取20个单株的叶片提取DNA,并利用MAS对目标代换片段标记和杂合标记进行筛选,根据标记带型选出纯合单片段代换系(S1-S4)和双片段代换系(D1-D5)。
对于2022年无杂合标记的单、双片段代换系和日本晴,在植株成熟后,选取各小区内中间2行3~7株共10个单株进行收割;对于该年含1个杂合标记的单、双片段代换系,2023年收取所有该标记纯合且与日本晴一致的株系。采用同样的方法测定9个F2代鉴定出QTL的农艺性状。
-
对任意1个SSSLi,提出假设H0:其代换片段不存在控制某一性状的QTL。对每个SSSL和受体亲本日本晴的某一性状在SPSS 25.0中进行ONE-WAY ANOVA和Duncan多重比较,当p<0.05时,表明假设不成立,即SSSLi的代换片段存在控制某一性状的QTL。在特定环境条件下,日本晴的遗传模型为:
单片段代换系的遗传模型为:
式中:Pi和P0分别表示SSSLi和日本晴的表型值;μ0为日本晴某一性状的表型值;ai为QTL的加性效应;ε为残差。
该QTL的加性效应(ai)为:
即认为表型差值的一半被估算为由遗传引起,所有计算均在Excel 2016中进行。
-
对任意1个DSSLij提出假设H0:位于“i”和“j”2个代换片段的QTL之间不存在上位性效应,即属于独立遗传,可表示为“2+0=1+1”。使用SPSS 25.0的TWO-WAY ANOVA和Duncan多重比较对日本晴、SSSLi、SSSLj和DSSLij的每一性状值进行双因素方差分析。当p<0.05时,则假设不成立。当Qi的主体间效应分析p<0.05时,表示i代换片段具有控制某一性状的QTL,当Qj的主体间效应分析p<0.05时,表示j代换片段具有控制某一性状的QTL,当Qi×Qj的主体间效应分析p>0.05时,接受H0,表明这两个QTL属于独立遗传;当p<0.05时,则拒绝H0,表明位于“i”和“j”代换片段的Qi和Qj间存在上位性效应,即“2+0≠1+1”。因此,
SSSLi的遗传模型为:
SSSLj的遗传模型为:
DSSLij的遗传模型为:
式中:Pi为SSSLi的表型值;Pj为SSSLj的表型值;ai和aj分别为代换片段i和j的QTL加性效应;Iij为代换片段i和j之间的上位性效应。上位性效应(I)为:
即其表型差值的一半被估算为由遗传引起。
1.1. 试验材料
1.2. 试验方法
1.2.1. 田间种植方法
1.2.2. 重要农艺性状考察
1.2.3. QTL定位
1.2.4. 次级单片段代换系和双片段代换系的培育
1.2.5. 基于单片段代换系的QTL验证和加性效应分析
1.2.6. 基于双片段代换系的上位性效应分析
-
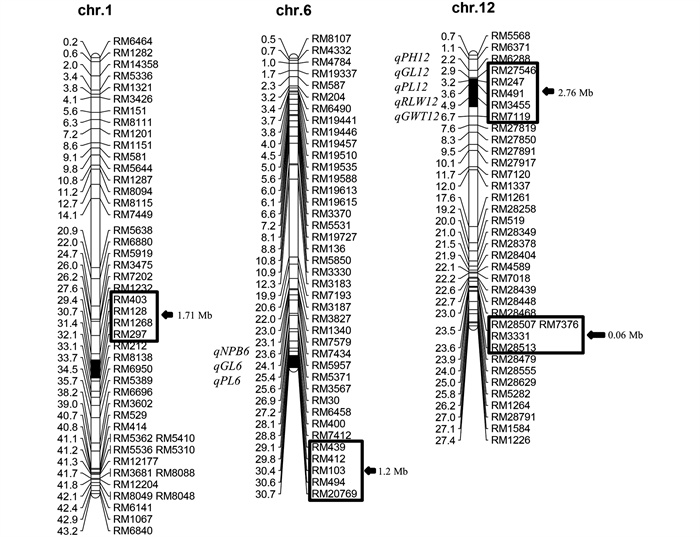
在前期选育得到含有4个代换片段Z454的基础上,本研究以代换片段上所有10个多态性SSR标记及代换片段外的18个SSR标记,用10株Z454植株对其代换片段和遗传背景纯度进行检测。结果表明,在所有植株中均含有4个一致的代换片段,且未检测到西恢18片段的其他残留片段。Z454的4个代换片段分别位于水稻的1、6和12号染色体上,总代换长度为5.73 Mb,其中最长的代换片段长度为2.76 Mb,最短的为0.06 Mb,平均为1.43 Mb(图 1)。
-
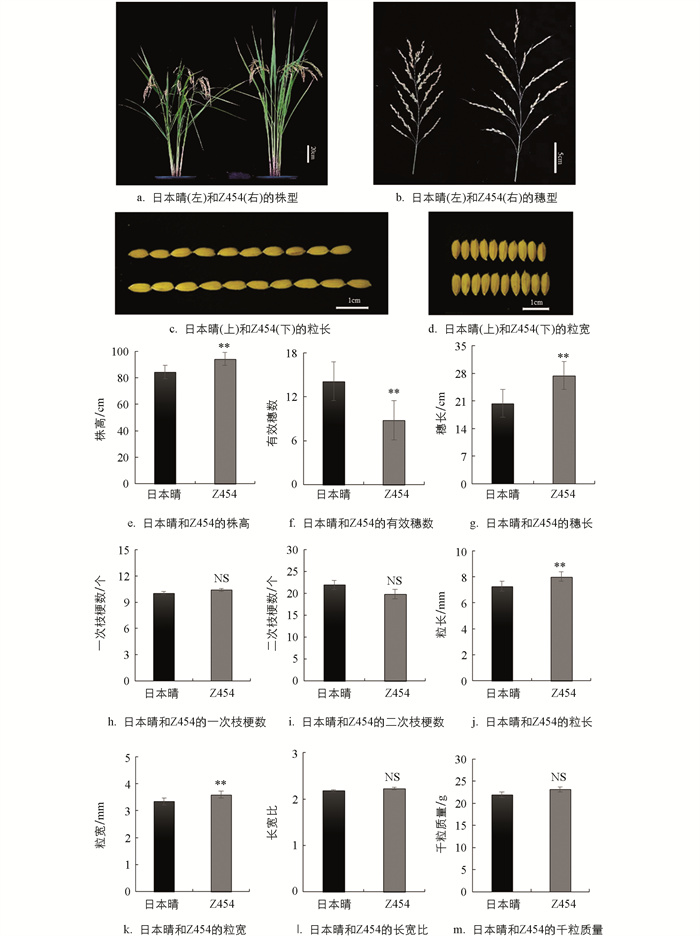
鉴于Z454与日本晴仅存在4个代换片段的差异,因此它们的株型整体相似(图 2a)。与日本晴相比,Z454的株高(94.14 cm)显著高于日本晴(84.32 cm),提高了11.65%(图 2a、2e)。Z454单株有效穗数(8.78)显著少于日本晴(14.11),减少了37.78%(图 2a、2f)。Z454的穗长(27.44 cm)比日本晴(20.38 cm)显著增加了34.64%(图 2b、2g)。Z454的粒长(8.01 mm)和粒宽(3.59 mm)比日本晴(7.28 mm、3.34 mm)分别增加了10.03%和7.48%,均达0.01的统计水平(图 2c、2d、2j、2k)。Z454与日本晴的一次枝梗数、二次枝梗数、长宽比和千粒质量差异均无统计学意义(图 2h、2i、2l、2m)。
-
以日本晴和Z454杂交构建的次级F2代作为QTL定位群体,共鉴定出8个水稻重要农艺性状的QTL,分布在第6和第12号染色体上,解释了表型变异的4.39%~32.68%(表 1)。这些QTL为1个控制株高的qPH12、1个控制一次枝梗数的qNPB6、2个控制穗长的qPL6和qPL12、2个控制粒长的qGL6和qGL12、1个控制长宽比的qRLW12和1个控制千粒质量的qGWT12。qNPB6、qPL6、qGL6均与第6号染色体的RM494紧密连锁,来自西恢18的等位基因的加性效应分别增加了0.40个一次枝梗数、0.82 cm的穗长和0.08 mm的粒长,分别解释了14.11%、11.89%和12.37%的表型变异。qPH12、qPL1 2、qGL12、qRLW12和qGWT12均与第12号染色体的RM247紧密连锁,来自西恢18的等位基因加性效应分别增加了0.76 cm的株高、0.95 cm的穗长、0.13 mm的粒长、0.03的长宽比和0.06 g的千粒质量,分别解释了4.39%、15.48%、32.68%、6.03%和5.65%的表型变异。
-
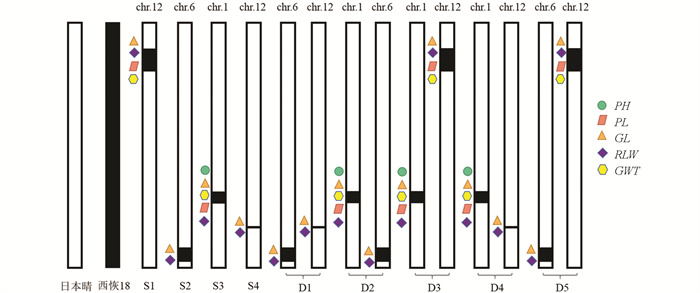
根据F2代的QTL定位结果,通过MAS在F3代中共构建了4个单片段代换系(SSSL,S1-S4)和5个双片段代换系(DSSL,D1-D5)。S1在第12号染色体上携带1个代换片段(RM27546-RM247-RM7119)。S2在第6号染色体上携带1个代换片段(RM439-RM494-RM20769)。S3在第1号染色体上携带1个代换片段(RM403-RM128-RM1268-RM297)。S4在第12号染色体上携带1个代换片段(RM28507-RM7376-RM3331-RM28513)。双片段代换系D1包含S2和S4的代换片段,D2包含S2和S3的代换片段,D3包含S1和S3的代换片段,D4包含S3和S4的代换片段,D5包含S1和S2的代换片段(图 3)。
-
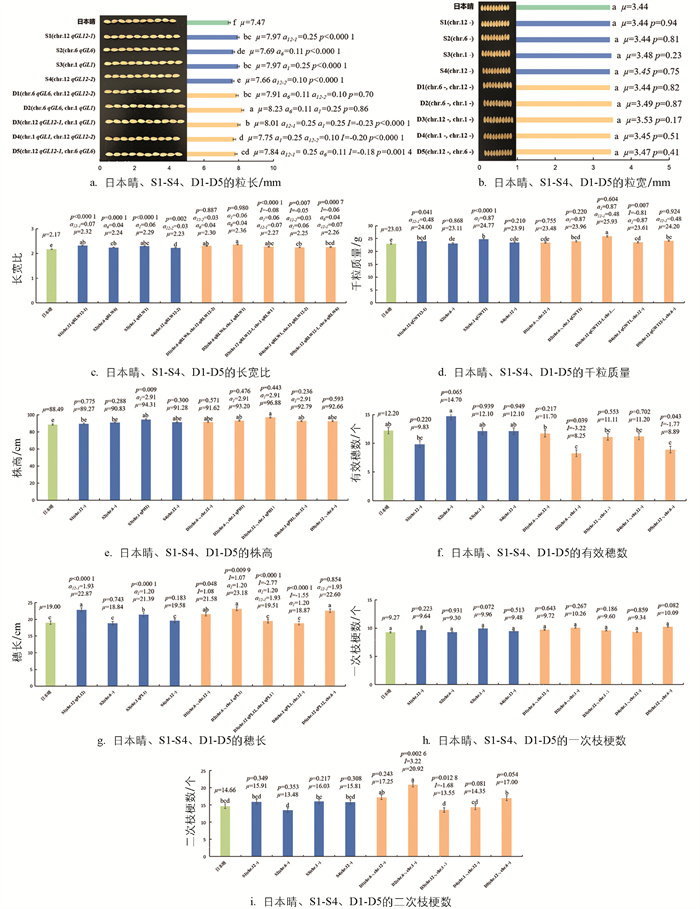
在单片段代换系S1-S4中,共鉴定到了13个重要农艺性状的QTL(图 4),包括4个粒长QTL(qGL12-1、qGL6、qGL1、qGL12-2),4个长宽比QTL(qRLW12-1、qRLW6、qRLW1、qRLW12-2),2个千粒质量QTL(qGWT12-1、qGWT1),1个株高QTL(qPH1),2个穗长QTL(qPL12-1、qPL1),其中5个QTL (qGL12-1、qPL12-1、qRLW12-1、qGWT12-1和qGL6)同时也能在F2代群体中检测到,表明其遗传受环境影响较小,年际间遗传稳定。qPH12、qNPB6、qPL6未能被相应的单片段代换系验证,暗示它们可能与环境的互作效应较大。此外,8个QTL(qRLW6、qPH1、qGL1、qGWT1、qPL1、qRLW1、qGL12-2、qRLW12-2)仅在单片段代换系中被检测到,表明SSSL具有比F2代更高的QTL检测效率。
含有qGL12-1 (a=0.25 mm)的S1、qGL6 (a=0.11 mm)的S2、qGL1 (a=0.25 mm)的S3和qGL12-2 (a=0.10 mm)的S4的粒长(7.97 mm、7.69 mm、7.97 mm和7.66 mm)均比日本晴的粒长(7.47 mm)显著增加(图 4a)。不含粒宽QTL的S1、S2、S3和S4的粒宽(3.44 mm、3.44 mm、3.48 mm、3.45 mm)与日本晴(3.44 mm)差异无统计学意义(图 4b)。分别携带qRLW12-1 (a=0.073)、qRLW6 (a=0.035)、qRLW1 (a=0.061)和qRLW12-2 (a=0.029)的S1、S2、S3和S4的长宽比(2.32、2.24、2.29、2.23)均比日本晴(2.17)显著增加(图 4c)。分别含有qGWT12-1 (a=0.48 g)和qGWT1 (a=0.87 g)的S1和S3的千粒质量(24.00 g和24.77 g)均比日本晴(23.03 g)显著增加,而不含该性状QTL的S2和S4的千粒质量(23.11 g、23.91 g)与日本晴差异无统计学意义(图 4d)。携带qPH1 (a=2.91 cm)的S3的株高(94.31 cm)比日本晴的株高(88.49 cm)显著增加,而不含该性状QTL的S1、S2和S4的株高(89.27 cm、90.83 cm、91.28 cm)与日本晴差异无统计学意义(图 4e)。不含有效穗数QTL的S1、S2、S3和S4的有效穗数(9.83个、14.7个、12.1个、12.1个)与日本晴(12.2个)差异无统计学意义(图 4f)。分别含有qPL12-1 (a=1.93 cm)和qPL1 (a=1.20 cm)的S1和S3的穗长(22.87 cm、21.39 cm)均比日本晴的穗长(19.00 cm)显著增加,而不含该性状QTL的S2和S4的穗长(18.84 cm、19.58 cm)与日本晴差异无统计学意义(图 4g)。无携带一次枝梗数QTL和二次枝梗数QTL的S1-S4与日本晴差异无统计学意义(图 4h、4i)。
-
粒长qGL6 (a=0.11 mm)与qGL12-2 (a=0.10 mm)在双片段代换系D1中独立遗传,则D1的粒长遗传效应为0.21 mm。此时,D1的粒长(7.91 mm)比含qGL12-2的S4和含qGL6的S2(7.66 cm、7.69 mm)显著增长(图 4a)。qGL6 (a=0.11 mm)与qGL1 (a=0.25 mm)在D2中也属独立遗传,因此D2的粒长遗传效应为0.36 mm。在表型上,D2的粒长(8.23 mm)均显著高于分别含有qGL6和qGL1的S2和S3(7.69 cm、7.97 mm)(图 4a)。qGL12-1 (a=0.25 mm)与qGL1 (a=0.25 mm)聚合后产生了-0.23 mm的上位性效应,使D3的粒长在遗传上增加了0.27 mm,与二者的加性效应相当,因此D3的粒长(8.01 mm)显著大于日本晴(7.47 mm),而与分别含有qGL12-1和qGL1的S1和S3(7.97 mm、7.97 mm)差异无统计学意义(图 4a)。qGL12-2 (a=0.10 mm)与qGL1 (a=0.25 mm)聚合后产生了-0.20 mm的上位性效应,据DSSL遗传模型,D4的粒长遗传效应为0.15 mm,因此D4的粒长(7.75 mm)正好介于含有qGL12-2的S4(7.66 mm)和含有qGL1的S3(7.97 mm)之间,比前者显著增长,而比后者显著缩短(图 4a)。qGL12-1 (a=0.25 mm)与qGL6 (a=0.11 mm)聚合后产生了-0.18 mm的上位性效应,使D5的粒长在遗传上增加了0.18 mm,因此D5的粒长(7.84 mm)也分别介于含有qGL12-1和qGL6的S1(7.97cm)和S2(7.69 mm)之间,但三者间差异无统计学意义(图 4a)。
长宽比qRLW6 (a=0.04)与qRLW12-2 (a=0.03)在D1中独立遗传,则D1的籽粒长宽比遗传效应为0.07,在表型上,D1的长宽比(2.30)显著大于日本晴(2.17)和含有qRLW12-2的S4(2.23),而比含有qRLW6的S2(2.24)虽有增加,但差异无统计学意义(图 4c)。qRLW6(a=0.04)与qRLW1(a=0.06)在D2中也属独立遗传,则D2的长宽比遗传效应为0.10;此时,D2的长宽比(2.36)显著大于日本晴(2.17)和含有qRLW6的S2(2.24),而与含有qRLW1的S3(2.29)差异无统计学意义(图 4c)。qRLW12-1 (a=0.07)与qRLW1 (a=0.06)聚合后产生了-0.08的上位性效应,使D3的长宽比在遗传上增加了0.05,因此D3的长宽比(2.27)小于含有qRLW12-1和qRLW1的S1(2.32)及S3(2.29),且差异无统计学意义(图 4c)。qRLW12-2 (a=0.03)与qRLW1 (a=0.06)聚合后产生了-0.05的上位性效应,使D4的长宽比在遗传上增加了0.04,导致D4的长宽比(2.25)虽比日本晴(2.17)显著增加,但与含有qRLW12-2的S4(2.23)和含有qRLW1的S3(2.29)差异无统计学意义,表型值介于二者之间(图 4c)。qRLW12-1 (a=0.07)与qRLW6 (a=0.04)聚合后产生了-0.06的上位性效应,使D5的长宽比在遗传上增加了0.05,导致D5的长宽比(2.26)显著大于日本晴(2.17),而与含有qRLW12-1的S1(2.32)和含有qRLW6的S2(2.24)差异无统计学意义(图 4c)。
千粒质量qGWT1(a=0.87 g)与无该性状QTL的第6号染色体代换片段独立遗传,则D2的千粒质量遗传效应为qGWT1的加性效应;相应地,D2的千粒质量(23.96 g)与含有qGWT1的S3(24.77 g)差异无统计学意义,而显著大于日本晴(23.03 g)(图 4d)。qGWT12-1(a=0.48 g)与qGWT1(a=0.87 g)在D3中独立遗传,因此D3的千粒质量遗传效应为1.35 g;相应地,D3的千粒质量(25.93 g)显著大于日本晴(23.03 g)、含有qGWT1的S3(24.77 g)和含有qGWT12-1的S1(24.00 g)(图 4d)。无千粒质量QTL的第12号染色体代换片段与qGWT1(a=0.87 g)聚合后产生了-0.81 g的上位性效应,则D4的千粒质量遗传效应为0.06 g;在表型上,D4的千粒质量(23.61 g)显著大于日本晴(23.03 g),显著小于含有qGWT1的S3(24.77 g),表明该代换片段影响了qGWT1的加性效应(图 4d)。qGWT12-1 (a=0.48 g)与无该性状QTL的第6号染色体代换片段在D5中独立遗传,则D5的千粒质量遗传效应为0.48 g,因此D5的千粒质量(24.20 g)与含有qGWT12-1的S1(24.00 g)差异无统计学意义,而显著大于日本晴(23.03 g)(图 4d)。D1的2个代换片段均无该性状的QTL,也无互作产生,在表型上,D1的千粒质量(23.48 g)与S2、S4和日本晴的千粒质量(23.11 g、23.91 g、23.03 g)差异无统计学意义(图 4d)。
无株高QTL的第6号染色体代换片段、第12号染色体的2个代换片段在D2、D3和D4中均不影响株高qPH1 (a=2.91 cm)的表达,因此D2、D3、D4的株高遗传效应均由qPH1的加性效应决定;相应地,三者的株高(93.20 cm、96.88 cm、92.79 cm)与含有qPH1的S3(94.31 cm)差异无统计学意义,且都显著大于日本晴的株高(88.49 cm)(图 4e)。在D1和D5中的2个代换片段均无株高QTL,也无互作的发生,D1和D5的株高(91.62 cm、92.66 cm)都与日本晴(88.49 cm)差异无统计学意义(图 4e)。
无有效穗数QTL的第1号染色体代换片段与无该性状QTL的第6号染色体代换片段聚合产生了-3.22个上位性效应,因此D2的遗传效应为每株减少了3.22个有效穗;相应地,D2的有效穗数(8.25个)显著少于日本晴、S3和S2的有效穗数(12.20个、12.10个和14.70个)。无有效穗数QTL的第12号染色体RM28507-RM7376-RM3331-RM28513代换片段与无该性状QTL的第6号染色体代换片段聚合产生了-1.77个有效穗的上位性效应,因此D5的遗传效应为-1.77个;在表型上,D5的有效穗数(8.89个)显著小于日本晴、S1和S2的有效穗数(12.20个、9.83个和14.70个)(图 4f)。D1、D3和D4的2个代换片段均无有效穗数QTL,也无互作的产生,因此三者的有效穗数(11.70个、11.11个、11.20个)均与日本晴的有效穗数(12.20个)差异无统计学意义(图 4)。
无穗长QTL的第6号染色体代换片段与第12号染色体代换片段在D1中聚合产生了1.08 cm的上位性效应,因此D1的遗传效应为1.08 cm;D1的穗长(21.58 cm)显著长于日本晴、S2和S4的穗长(19.00 cm、18.84 cm和19.58 cm)(图 4g)。无穗长QTL的第6号染色体代换片段与qPL1(a=1.20 cm)聚合产生了1.07 cm的上位性效应,因此D2的遗传效应为2.27 cm;D2的穗长(23.18 cm)显著长于日本晴(19.00 cm)、含有第6号染色体代换片段的S2(18.84 cm)和含有qPL1的S3(21.39 cm)(图 4g)。qPL12-1(a=1.93 cm)与qPL1(a=1.20 cm)聚合后产生了-2.77 cm的上位性效应,因此D3的穗长遗传效应为0.36 cm;在表型上,D3的穗长(19.51 cm)与日本晴(19.00 cm)差异无统计学意义,而显著短于含有qPL12-1的S1(22.87 cm)和含有qPL1的S3(21.39 cm)(图 4g)。无穗长QTL的第12号染色体代换片段与qPL1(a=1.20 cm)在D4中聚合产生了-1.55 cm的上位性效应,使D4的穗长在遗传上减少了0.35 cm;相应地,D4的穗长(18.87 cm)与日本晴(19.00 cm)和含有第12号染色体RM28507-RM7376-RM3331-RM28513代换片段的S4(19.58 cm)差异无统计学意义,而显著短于含有qPL1的S3(21.39 cm)(图 4g)。qPL12-1(a=1.93 cm)与无穗长QTL的第6号染色体代换片段独立遗传,因此D5的遗传效应由qPL12-1的加性效应决定;在表型上,D5的穗长(22.60 cm)显著长于日本晴(19.00 cm)和含有第6号染色体代换片段的S2(18.84 cm),而与含有qPL12-1的S1(22.87 cm)差异无统计学意义(图 4g)。
无二次枝梗数QTL的第1染色体代换片段与第6号染色体代换片段聚合产生了3.22个二次枝梗数的上位性效应,则D2的遗传效应为每穗增加了3.22个;相应地,D2的二次枝梗数(20.92个)显著大于日本晴、S3和S2(14.66个、16.03个和13.48个)(图 4i)。无二次枝梗数QTL的第12号染色体代换片段与第1染色体代换片段在D3中聚合产生了-1.68个二次枝梗数的上位性效应,因此D3的遗传效应为每穗减少了1.68个二次枝梗数;在表型上,D3的二次枝梗数(13.55个)显著小于含有第1染色体代换片段的S3(16.03个),而比日本晴(14.66个)和含有第12号染色体代换片段的S1(15.91个)虽有所减少,但差异无统计学意义(图 4i)。D1、D4和D5的2个代换片段无该性状的QTL,也无互作产生,三者的二次枝梗数(17.25个、14.35个和17.00个)与日本晴(14.66个)差异无统计学意义(图 4i)。
此外,对于粒宽与一次枝梗数2个性状,双片段代换系D1、D2、D3、D4和D5的任何2个代换片段均无该性状的QTL,也无互作发生,这些双片段代换系与日本晴的粒宽和一次枝梗数差异均无统计学意义(图 4b、4h)。
2.1. CSSL-Z454代换片段的鉴定
2.2. Z454和日本晴的表型分析
2.3. Z454的代换片段上携带的水稻重要农艺性状QTL
2.4. 次级单、双片段代换系的选育
2.5. 基于单片段代换系(S1-S4)的9个性状QTL鉴定与加性效应分析
2.6. 基于双片段代换系(D1-D5)的9个性状非等位QTL间的上位性效应分析
-
随着科学技术的发展,育种方法逐渐由传统的选择育种、杂交育种、标记辅助选择育种到如今的分子设计育种[21]。分子设计育种相较于前3种方法,不仅需要明确各基因的遗传效应,更需要清晰多个基因之间的互作情况,在此基础上才能够通过计算机的选择实现直接定向育种[22]。水稻的产量属于由多个微小基因控制的复杂数量性状,需要通过CSSL等作图群体将数量性状分解为单个孟德尔因子[16]。单片段代换系(SSSL)的构建不仅可使QTL定位更加准确,同时也为分子设计育种奠定了良好的基础。如Zhang[21]通过HJX74-SSSL文库的构建,实现了多个优良等位基因的快速、精准聚合,实现了多个水稻新品种的培育。然而,单片段代换系的构建过程复杂,需多年多代才能完成,在本研究中,我们首先鉴定了1个以日本晴作为受体亲本、西恢18作为供体亲本的4个代换片段CSSL-Z454,然后通过日本晴/Z454的F2代经重要性状的QTL和分子标记辅助选择,最后构建了含重要性状QTL的4个单片段代换系(S1-S4)和5个双片段代换系(D1-D5)。这些SSSL不仅验证了F2代鉴定的5个QTL(qGL12-1、qPL12-1、qRLW12-1、qGWT12-1和qGL6),还检测到8个新的QTL(qRLW6、qPH1、qGL1、qGWT1、qPL1、qRLW1、qGL12-2、qRLW12-2),表明单片段代换系由于遗传背景与受体亲本的高度一致性,极大地提高了QTL鉴定的精准性和检测效率。此前的许多研究已证实SSSL具有更高的QTL检测效率[23-24]。携带qGL12-1、qGWT12-1、qPL12-1的单片段代换系S1粒长(7.97 mm)、千粒质量(24.00 g)和穗长(22.87 cm)比受体日本晴显著增加,而其余性状差异不大;含有qGL1、qGWT1、qPH1、qPL1的S3粒长(7.97 mm)、千粒质量(24.77 g)、株高(94.31 cm)和穗长(21.39 cm)均比日本晴显著增加,而与其余性状间差异不大;含qGL6的S2和含qGL12-2的S4的粒长(7.69 cm和7.66 mm)比日本晴显著增加,而与其他性状间差异无统计学意义。综上所述,单片段代换系S1-S4均具有不同育种的有利性状QTL,这保证了以SSSL进行目标性状改良的高效性和准确性,因此SSSL为平台的高产分子设计育种提供了可靠的遗传基因和育种资源。
-
通过日本晴/Z454的F2代和进一步分解构建的SSSL,我们共鉴定到13个重要性状QTL。通过与前人在相应区间内报道的基因相比,DYW3和OsGATA6位于qGL1、qGLW、qGTW1、qPH1和qPL1的代换片段内,这些QTL均与分子标记RM128(30.7 Mb)紧密连锁,DYW3(30.77 Mb)与该标记相距0.07 Mb,OsGATA6(31.17 Mb)与该标记相距1.1 Mb。DYW3能够与FLO22互作,可能协同参与线粒体编辑。DYW3功能的缺失会对水稻植株生长和胚乳发育造成影响,使株高降低、千粒质量下降[25]。据功能分析,DYW3可作为本研究鉴定的qGTW1和qPH1的候选基因。OsGATA6能够负调控水稻抽穗期,正调控穗发育,并影响籽粒大小。在过表达OsGATA6的植株中出现抽穗延迟,一、二次枝梗数增加,每穗粒数增加,但籽粒整体变小,粒长、粒宽和千粒质量均下降的表型。OsGATA6主要通过影响细胞分裂来调节籽粒大小[26]。据功能分析,OsGATA6可作为本研究鉴定qGL1、qGLW、qGTW1、qPL1的候选基因。qGL6和qRLW6与分子标记RM494(30.60 Mb)连锁,在此区间内发现1个与粒型相关的基因OsPRA2。OsPRA2(30.33 Mb)与该标记相距0.27 Mb,该基因的过表达会使水稻株系出现株高下降、叶倾角变小、种子整体变小和节间伸长模式异常的表型,同时对外源油菜素内酯敏感性降低,一些BR合成基因的表达出现上调。通过RNAi抑制OsPRA2表达会导致植株表现出相反的表型[27],OsPRA2可作为鉴定qGL6和qRLW6的候选基因。qGL12-1、qRLW12-1、qGWT12-1和qPL12-1与分子标记RM247(3.19 Mb)紧密连锁,在此区间发现1个与粒型相关的基因OsYUC11。OsYUC11(4.51 Mb)与该标记相距1.32 Mb,该基因的激活与表达能够促进生长素生物合成,生长素水平的提高刺激了胚乳细胞中蔗糖代谢和淀粉合成相关基因的表达,进而增加了淀粉积累以及籽粒大小[28]。OsYUC11可作为鉴定qGL12-1、qRLW12-1、qGWT12-1和qPL12-1的候选基因,穗长的变化可能是该基因存在一因多效的现象。qGL12-2、qRLW12-2被定位于RM28507(23.5 Mb)-RM7376-RM3331-RM28513(23.6 Mb)区间范围内,该区间包含LOC_Os12g38290、LOC_Os12g38300、LOC_Os12g38310、LOC_Os12g38320、LOC_Os12g38330、LOC_Os12g3834、LOC_Os12g38350、LOC_Os12g38360、LOC_Os12g38370、LOC_Os12g38380、LOC_Os12g38400等11个基因,其中,LOC_Os12g38400与水稻抗逆性密切相关[29],其余基因无研究报道,说明qGL12-2、qRLW12-2可能是本研究新鉴定的QTL,后续将对范围内的基因进行进一步DNA测序。
-
水稻产量性状属于典型的数量性状,在由许多微效基因控制的同时,多个QTL之间还存在着互作。Balakrishnan等[30]认为,对于复杂性状,QTL间的上位性效应是性状表现的重要遗传成分。存在互作的QTL数量往往大于或等于加性QTL的数量,同时其大小也有灵活的变化范围。对自然遗传变异而言,上位性效应是重要的组成部分[31],在最近的研究中,Cyplik等[32]进一步提出QTL-QTL-QTL三重相互作用对数量性状表达有显著的影响,同一性状不同QTL在不同材料中的遗传也表现出极大的多样性[23, 33-35]。在本研究中,以日本晴/Z454的F2代和进一步分解构建的SSSL与DSSL进行了多个QTL组合的上位性检验,结果表明,同一QTL与不同QTL聚合会有不同的遗传模式,如粒长qGL6和qGL12-2与qGL1在D1和D2中均独立遗传,相应产生的遗传效应(0.21 mm和0.36 mm)均明显大于各自单个QTL的加性效应,因此D1(7.91 mm)显著长于含qGL12-2的S4和含qGL6的S2(7.66 cm和7.69 mm),D2的粒长(8.23 mm)显著长于含有qGL1的S3(7.97 mm)和含有qGL6的S2(7.69 cm),表明要想得到更长的籽粒,可选择qGL6与qGL12-2或qGL1聚合。而qGL6与qGL12-1聚合后产生了-0.18 mm的上位性效应,致使D5的粒长遗传效应0.18 mm与二者的加性效应相当,因此D5的粒长(7.84 mm)与含有qGL12-1和qGL6的S1和S2粒长(7.97 cm和7.69 mm)差异无统计学意义,表明qGL6与qGL12-1聚合对改进粒长性状无实际意义。qGL12-2与qGL1聚合后产生了-0.20 mm的上位性效应,致使D4的遗传效应0.15 mm介于单个QTL的加性效应之间,因此D4的粒长(7.75 mm)显著长于含有qGL12-2的S4(7.66 mm),而显著短于含有qGL1的S3(7.97 mm),因此,要想得到中间粒长,可选择这两个QTL聚合。qGL12-1与qGL1聚合也产生了-0.23 mm的上位性效应,致使D3的遗传效应0.27 mm与二者的加性效应相当,因此,D3的粒长(8.01 mm)与分别含有qGL12-1和qGL1的S1及S3(7.97 cm和7.97 mm)差异无统计学意义,选择这两个QTL聚合在改进粒长上无实际意义。由此可见,明确每个QTL的加性效应及与不同QTL间的遗传模式,对分子设计育种选择合适的QTL有重要的应用价值。
既然每个SSSL还含有不同性状的QTL,而且这些性状间还存在不同程度的相关或无关[36-38],因此要想达到最佳目标性状的效果,需综合分析DSSL所含不同性状QTL间的遗传模式。鉴于D1的粒长qGL6和qGL12-2独立遗传,二者聚合产生0.21 mm的粒长遗传效应,使得D1的粒长(7.91 mm)显著增加,达到改良粒长的效果;D1的穗长(21.58 cm)比受体日本晴和相应的单片段代换系S2和S4穗长显著增加;而D1的粒宽(3.44 mm)、千粒质量(23.48 g)、株高(91.62 cm)、有效穗数(11.70个)、一、二次枝梗数(9.72个和17.25个)与相应的单片段代换系S2和S4及受体日本晴差异无统计学意义,因此利用D1或S2和S4杂交作为育种材料能够实现其余性状不变的情况下迅速改进粒长和穗长的育种目标。D2的粒长qGL6和qGL1聚合产生的遗传效应为0.36 mm,明显大于qGL6和qGL1的加性效应(0.11 cm和0.25 mm),D2的粒长(8.23 mm)显著长于日本晴和相应的单片段代换系S2和S3;D2的穗长(23.18 cm)和二次枝梗数(20.92个)也均显著大于日本晴、S2和S3;然而D2的2个代换片段间产生了-3.22个有效穗数的上位性效应,致使D2的有效穗数(8.25个)显著少于日本晴、S2和S3,因此以D2或S2与S3杂交为育种材料可在除有效穗减少外实现长粒、长穗的育种目标。D3的粒长qGL12-1和qGL1聚合产生0.27 mm的遗传效应,与二者的加性效应(0.25 mm和0.25 mm)相当,其粒长(8.01 mm)与相应的单片段代换系S1和S3差异无统计学意义;D3的千粒质量qGWT12-1和qGWT1聚合产生1.35 g的遗传效应,使D3的千粒质量(25.93 g)比相应单片段代换系S1和S3显著增加;D3的穗长qPL12-1和qPL1聚合产生的遗传效应为0.36 cm,导致D3的穗长(19.51 cm)与日本晴差异无统计学意义,而显著小于S1和S3;D3的株高(96.88 cm)与含qPH1的S3差异无统计学意义,但比日本晴的株高(88.49 cm)显著增加;D3的粒宽、有效穗数、一、二次枝梗数与日本晴差异无统计学意义,因此以D3或S1与S3杂交为育种材料可在多数性状不变下迅速改良粒长、粒质量、穗长和株高等性状。D4的粒长qGL1和qGL12-2聚合产生的遗传效应为0.15 mm,介于qGL1和qGL12-2的加性效应之间,导致D4的粒长(7.75 mm)显著大于日本晴,短于S3(7.97 mm)而长于S4(7.66 mm);D4的株高(92.79 cm)也与含qPH1的S3差异无统计学意义,但显著高于日本晴。D4的其余性状与日本晴和S3和S4间差异无统计学意义,因此以D4为育种材料或选择S3和S4杂交与S3的粒长和株高QTL遗传效应相当,没实际意义,在设计育种中,选择S3进行改良株高和粒长即可。D5的粒长qGL12-1和qGL6聚合产生0.18 mm的遗传效应,介于qGL12-1和qGL6的加性效应(0.25 cm和0.11 mm)之间,导致D5的粒长(7.84 mm)与S1和S2差异无统计学意义;D5的穗长(22.60 cm)和千粒质量(24.20 g)与含有qPL12-1和qGWT12-1的S1差异无统计学意义;此外,D5的株高、一、二次枝梗数、粒宽等与日本晴和S1及S2差异无统计学意义,因此,以D5或S1和S2杂交为育种材料与S1无明显差异,在设计育种中,选择S1改良粒长、穗长和粒质量即可。由此可见,与传统育种相比,以SSSL为平台,可实现在非目标性状不变的情况下,目标QTL的精准聚合,并可较准确地预测改良性状的表型,实现快速、高效地设计育种。
-
为构建一套覆盖水稻全基因组的粳型亲籼水稻单片段代换系文库,本研究以一个日本晴为遗传背景来自籼型恢复系西恢18的4个代换片段的CSSL-Z454为研究材料,通过QTL定位,进一步构建了4个单片段代换系和5个双片段代换系,共鉴定出13个水稻重要性状的QTL,包括4个粒长QTL(qGL12-1、qGL6、qGL1、qGL12-2),4个长宽比QTL(qRLW12-1、qRLW6、qRLW1、qRLW12-2),2个千粒质量QTL(qGWT12-1、qGWT1),1个株高QTL(qPH1),2个穗长QTL(qPL12-1、qPL1),其中,qGL12-2、qRLW12-2可能是本研究新发现的QTL。单片段代换系S1-S4均具有不同育种的有利性状QTL,如S1含增加0.25 mm的粒长qGL12-1、增加千粒质量0.48 g的qGWT12-1和增加穗长1.93 cm的qPL12-1等,而其余性状与受体差异无统计学意义,这保证了以SSSL进行目标性状改良的高效性和准确性。此外,经9个水稻重要性状QTL的加性和上位性效应分析表明,同一QTL与不同QTL互作会产生不同的遗传模式,如粒长qGL6与qGL12-2及与qGL1在D1和D2中均独立遗传,而qGL6与qGL12-1在D5中聚合产生了-0.18 mm的上位性效应;qGL12-2与qGL1在D4中聚合产生了-0.20 mm的上位性效应;qGL12-1与qGL1在D3中聚合也产生了-0.23 mm的上位性效应,因此以SSSL及相应DSSL鉴定每个QTL及QTL间的遗传模式对设计育种非常必要。经过综合性分析,本研究发现用D1或S2和S4杂交作为育种材料能够实现其余性状不变的情况下迅速改进粒长和穗长的育种目标;以D2或S2与S3杂交为育种材料可在除有效穗减少外实现长粒、长穗的育种目标;以D3或S1与S3杂交为育种材料可在多数性状不变的情况下改良粒长、粒质量、穗长和株高性状;D4和D5作为育种材料的效果与S3和S1相当,因此可直接用S3和S1改良粒长性状。本研究为SSSL平台的水稻设计育种选择合适QTL和SSSL或DSSL提供了可靠的理论依据和育种资源。