



 下载:
下载:




-
开放科学(资源服务)标识码(OSID):

-
蜂巢小甲虫(Aethina tumida)属于鞘翅目露尾甲科, 是一种重要的入侵害虫, 对蜂产业的可持续发展带来重大威胁, 同时也对水果产业造成显著的经济损失。该害虫源于非洲, 1998年首次发现于美国的佛罗里达州, 后传播到埃及、韩国、毛里求斯等国家[1-4]。2017年, 首次发现于我国广东省, 随后扩散到海南省等地的蜜蜂养殖场[5-6]。蜂巢小甲虫是一种杂食性害虫, 通过取食蜂蜜、花粉、蜂卵以及幼虫等方式为害蜂群, 其幼虫排泄物污染蜂蜜, 导致蜂蜜失去食用价值, 严重时可使蜂群在两周内因蜜蜂逃逸而崩溃。此外, 蜂巢小甲虫寄主范围广泛, 还可通过取食为害香蕉、哈密瓜、樱桃等果实, 给水果产业造成严重经济损失[7]。目前, 蜂巢小甲虫的防控手段较为单一, 主要依赖物理和化学的手段进行防治。但物理防治效果不佳, 而化学防治虽高效, 但存在污染蜂蜜、伤害蜜蜂等风险。因此, 开发绿色高效, 对环境友好的防控策略势在必行。基于寄主挥发物的引诱剂或驱避剂具有环保、减少农药使用量、不伤害非靶标昆虫, 且害虫不易产生抗性的优点, 为防控蜂巢小甲虫提供了一种安全、绿色高效的新途径。
气味结合蛋白(odorant binding proteins, OBPs)是昆虫嗅觉系统中的关键作用分子之一, 以高浓度分布在嗅觉感器中, 能够识别并结合疏水性的挥发物, 并将其转运到嗅觉受体, 触发嗅觉信号传导[8]。挥发物进入嗅觉感器后, 首先与OBPs识别结合, 再通过转运作用穿过水相的淋巴液抵达受体, 这个过程被认为是昆虫识别挥发物的首步生理反应, 对于昆虫与外界进行化学交流有重要意义[9]。随着分子生物学技术的发展, 试验验证表明, OBPs通过选择性识别与转运挥发物实现其嗅觉功能。利用该机制可反向筛选调控昆虫嗅觉行为的挥发物, 为开发新型害虫引诱剂或驱避剂提供了新思路, 但是研究进展缓慢[10]。究其原因, 主要与传统的研究方法存在较大不足有关, 当前研究OBPs与气味分子作用的传统方法是荧光竞争结合试验, 该方法需要表达、纯化目标OBPs, 费用高、耗时长、局限性强, 严重制约着筛选有效挥发物的进程。
分子对接是基于Fischer在1894年提出的“锁钥学说”和Koshland在1958年提出的“诱导契合”两个学说的基础上, 利用模型模拟, 研究蛋白质与受体间相互作用的方法。通过对蛋白质进行同源建模以及对构建模型进行评价, 筛选出最佳的蛋白质模型与受体进行分子对接, 寻找结合能力最强的蛋白质-受体复合物。在分子对接过程中, 将三维结构数据库中的受体逐一与靶标蛋白质的活性位点匹配, 通过不断优化蛋白质的位置、构象等, 寻找受体与靶标蛋白质作用的最佳构象[11]。与常规的荧光竞争结合试验相比, 分子对接无需纯化OBPs, 具有费用低、效率高, 可快速大批量筛选出具有调控害虫嗅觉行为的挥发物, 因此应用前景广阔。
目前, 分子对接技术已成功应用于膜翅目、鞘翅目、半翅目等昆虫的OBPs与挥发物作用机制的研究中[12-15]。本研究以蜂巢小甲虫触角特异性高表达的OBP3为研究对象, 利用前期所测转录组数据, 选取从蜂巢材料及工蜂中鉴定出来的46种挥发物[16-17], 通过同源建模和分子对接技术, 筛选与OBP3结合能力强的挥发物, 以揭示OBP3在嗅觉中的识别作用, 为开发高效的蜂巢小甲虫引诱剂或驱避剂提供理论依据。
全文HTML
-
序列来源: 基于本课题组前期对蜂巢小甲虫成虫进行转录组测序所得到的OBP序列数据, 选取在触角特异性高表达的OBP3作为研究对象。
挥发物: 选取从蜂巢材料和工蜂中提取出来的46种挥发物用于分子对接。
-
使用SignalP 6.0(
https://services.healthtech.dtu.dk/services/SignalP-6.0 )预测OBP3氨基酸序列的信号肽位置, 去除信号肽序列后进行同源建模。通过Swiss-Model在线软件(https://swissmodel.expasy.org )预测构建三维结构的最佳模型, 综合GMQE、QMEAN以及模型氨基酸序列与OBP3序列的相似度和覆盖率等参数进行综合评价后, 选取最佳模型。用Clustal X软件比较OBP3和模型蛋白质氨基酸序列的相似度, 并通过Webologo 3(https://weblogo.threeplusone.com )将结果进行可视化处理。 -
为确保同源建模结果可用于分子对接, 利用SAVEs 6.0(
https://saves.mbi.ucla.edu )对建模结果合理性进行评价。利用Procheck对蛋白质几何形状进行评估, 以评价蛋白质的立体参数: 通过分析对氨基酸残基碳骨架的二面角在蛋白质或多肽链中的分布情况, 生成拉氏构象图。将结果用不同颜色的曲线划分为4个部分, 分别是最合理区(the most favoured regions)、其他合理区(the additional allowed regions)、一般合理区(the generously allowed regions)、不合理区(the disallowed regions)。从拉氏构象图中可获得各个氨基酸残基的二面角分布情况, 当90%以上的残基分布于最合理区时, 则认为蛋白质的结构合理。Verify 3D对同源建模所得模型进行可视化分析, 以判断模型与其本身氨基酸序列之间的兼容性。当80%以上的氨基酸残基得分大于0.2时, 即判定目标蛋白质模型结构为高质量。ERRAT计算3.5 Å范围之内, 不同原子之间形成的非键相互作用的数目, 结果应大于50%。若同源建模结果均满足上述条件, 则认为所建模型可用于分子对接。 -
配体结构文件: 从ChemSpider网站(
https://www.chemspider.com/StructureSearch.aspx )下载所选挥发物三维结构的SDF文件, 利用Open-Babel 3.0对其进行结构转换, 储存为PDB格式文件, 再用AutoDock tool 1.5.6软件进行结构优化, 保存为PDBQT格式文件。受体结构文件: 采用AutoDock Tool 1.5.6软件对评价合格的模型进行去水加氢处理, 储存为PDBQT格式文件。
分子对接分析采用AutoDock Tool 1.5.6软件进行半柔性对接, 在Grid Box中设置受体与配体的结合空间, 坐标为x=-2.695, y=6.583, z=-15.535, 格点间距为0.375 nm, 使用拉马克遗传算法计算结合自由能, 选择结合能力最强(即结合能最低)的构象[18], 利用Discovery Studio软件对结果进行可视化。
1.1. 试验材料
1.2. 试验方法
1.2.1. OBP3的同源建模
1.2.2. OBP3同源建模结果评估
1.2.3. OBP3与挥发物的分子对接分析
-
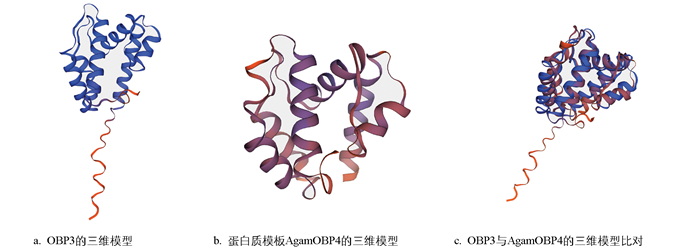
本研究聚焦蜂巢小甲虫气味结合蛋白OBP3的分子特征与结构建模。该蛋白基因编码148个氨基酸, 系统发育分析显示其N端存在16个氨基酸组成的信号肽。通过Swiss-Model对其进行同源模板搜索, 发现冈比亚按蚊(Anopheles gambiae)的AgamOBP4(模板编号为8c6e.1.A)具有显著的结构同源性: 氨基酸覆盖率为89%, 其中6个特征性半胱氨酸残基的空间排布完全保守(图 1)。尽管两者整体序列相似度31%, 但其结构特征符合昆虫OBPs家族典型的结构保守性, 满足同源建模的要求。因此, 以AgamOBP4为模板构建OBP3的三维结构模型(图 2)。
-
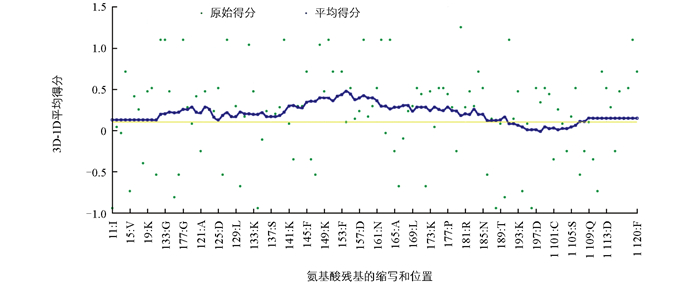
模型质量评估结果表明, OBP3结构高度合理。Procheck结果显示, 92.2%的氨基酸位于最合理区, 7.8%位于其他合理区, 无氨基酸分布于一般合理区以及不合理区(图 3)。Verify 3D结果表明, 86.67%的氨基酸残基的3D/1D值得分大于0.2(图 4), 反映出该模型的一级序列与三维结构有较高的相容性, 序列与结构之间匹配度较高。ERRAT值为98.2%(图 5), 远大于50%, 说明构建的模型中非键合相互作用合理。综合以上评估, 所构建的OBP3三维结构模型质量可靠, 适用于后续分子对接。
-
分子对接分析表明OBP3与46种挥发物的结合能的差异显著(表 1)。其中, OBP3与2-乙基己基-6-乙基-3-辛烷基邻苯二甲酸酯的结合能最低(-12.15 kcal/mol), 显示出最强的结合能力; 与3, 5, 24-trimethyltetracontane的结合能最高(0.76 kcal/mol), 结合能力最弱。为进一步明确OBP3的生物学功能, 选取Dekebo等[19]报道的5种对蜂巢小甲虫具有行为反应的挥发物进行详细分析, 包括对雌雄都有强烈的引诱作用的(Z, Z, Z)-9, 12, 15-十八碳三烯酸乙基酯和十六烷酸乙酯, 以及具有驱避作用的二十四烷、(Z)-9-十八碳烯酰胺和5-甲基-2-苯基-1H-吲哚。
-
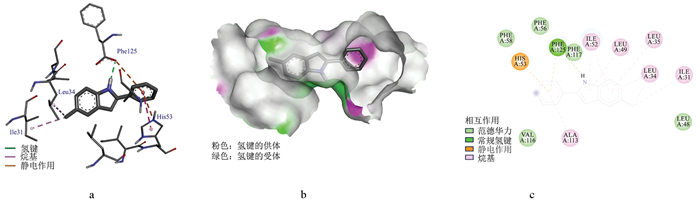
OBP3与(Z, Z, Z)-9, 12, 15-十八碳三烯酸乙基酯的结合能为-6.34 kcal/mol, 表明两者具有较强的结合特性(图 6)。三维结构图和口袋模型(图 6a、b)显示, 该小分子嵌入OBP3的结合空腔内, 二维作用图(图 6c)显示二者的结合主要通过疏水作用和范德华力实现。其中, 范德华力涉及氨基酸Ala70、Phe58、Ala113、Phe117、Phe125、Leu35、Leu65、His53、Leu34、Leu49、Ile31、Leu48; 疏水作用涉及氨基酸Val13、Ile52、Ile16、Phe56、Ile112、Val116、Ala92、Cys95、Ile91。
-
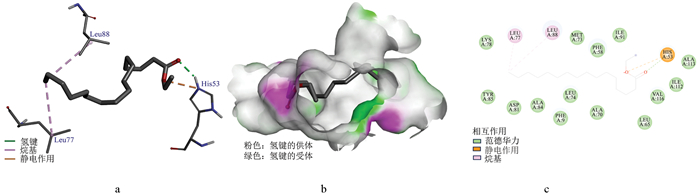
OBP3与十六烷酸乙酯的分子对接图见图 7, 二者结合能为-5.48 kcal/mol。基于OBP3的三维结构解析与结合口袋建模分析, 该小分子被准确定位于其配体结合腔内部(图 7a、b), 其主要通过疏水作用、范德华力、氢键及静电作用结合(图 7c)。疏水作用的氨基酸位点为Leu77和Leu88; 范德华力作用位点为Lys78、Tyr85、Asp81、Ala84、Phe9、Leu74、Ala70、Leu65、Val116、Ile112、Ala113、Ile91、Phe58、Met73; 氢键和静电作用位点均为氨基酸His53。
-
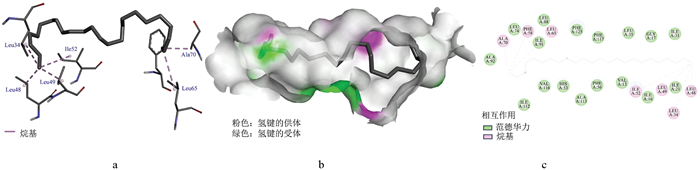
OBP3与二十四烷的分子对接图见图 8, 二者结合能为-5.70 kcal/mol, 二十四烷紧密嵌合于OBP3的结合空腔内, 周围分子作用增强结构稳定性(图 8a、b)。疏水作用涉及的氨基酸有Ala70、Phe58、Leu65、Ile52、Leu49、Leu34、Leu48; 范德华力作用位点涉及氨基酸Leu74、Leu88、Ile91、Ala92、Phe125、Phe117、Leu35、Gly17、Ile31、Ile21、Ile16、Val13、Phe56、Ala113、His53、Val116、Ile112(图 8c)。
-
OBP3与(Z)-9-十八碳烯酰胺的分子对接图见图 9, 二者结合能为-5.53 kcal/mol, 小分子通过范德华力、疏水作用和氢键稳定嵌入OBP3的活性口袋中(图 9b、c)。疏水作用位点涉及氨基酸有Ala92、Cys95、Val116、Ile112、Leu65; 范德华力作用涉及的氨基酸有Ile91、His53、Ala113、Phe56、Phe117、Phe125、Ile52、Leu49、Ile21、Leu48、Gly32、Leu35、Leu34、Phe58、Leu74、Leu88、Ala70; 氢键由氨基酸Ile31形成。
-
OBP3与5-甲基-2-苯基-1H-吲哚的分子对接图见图 10, 分子对接模拟显示该小分子以低结合自由能精准嵌入OBP3的结合口袋(图 10a、b), 二者结合能为-6.80 kcal/mol, 表明OBP3与5-甲基-2-苯基-1H-吲哚有较强的结合力。二者之间除了通过疏水作用、范德华力和静电作用, 还形成了氢键共同维持其结构的稳定性(图 10c)。疏水作用位点涉及氨基酸Ile52、Leu49、Leu35、Leu34、Ile31、Ala113; 范德华力作用位点涉及氨基酸Phe58、Phe56、Leu48、Val116; 静电作用位点涉及氨基酸His53和Phe125; 氢键作用位点由氨基酸Phe117形成。
2.1. OBP3同源建模结果
2.2. OBP3同源建模结果的评估
2.3. OBP3分子对接结果
2.3.1. OBP3与46种挥发性化合物的结合情况
2.3.2. OBP3与(Z, Z, Z)-9, 12, 15-十八碳三烯酸乙基酯的结合评估
2.3.3. OBP3与十六烷酸乙酯的结合评估
2.3.4. OBP3与二十四烷的结合评估
2.3.5. OBP3与(Z)-9-十八碳烯酰胺的结合评估
2.3.6. OBP3与5-甲基-2-苯基-1H-吲哚的结合评估
-
昆虫依赖其灵敏的嗅觉系统从复杂的环境中获得特异的化学信息, 激发相应的神经生理冲动, 引发相应的行为, 实现对寄主、配偶和产卵地点的准确定位和评估[20-21]。气味结合蛋白在这个过程中起关键作用, 它们识别并结合环境中的疏水性挥发物, 并将其转运到嗅觉受体[22]。传统的荧光竞争结合试验虽可验证OBPs与配体的结合特性, 但因其蛋白表达耗时长、成本高而受限。相比之下, 分子对接技术通过模拟配体与受体的相互作用, 有效弥补了这一不足, 成为近年来研究OBPs与挥发物作用机制及配体筛选的重要工具, 其优势在多种昆虫的研究中得到了充分的验证。例如, Li等[23]利用分子对接技术, 分析了桑螟(Glyphodes pyloalis)触角中高表达的GOBP1和GOBP2与植物挥发物的结合模式, 并结合定点突变技术, 揭示了GOBPs如何识别并运输受害植物挥发物, 进而调控桑螟雄蛾的嗅觉行为反应, 为分子对接技术在嗅觉机制研究中的应用提供了有力支持。Jing等[24]通过分子对接技术, 精准解析了松毛虫(Conogethes pinicolalis)气味结合蛋白GOBP1的多重配体识别机制, 证明其在寄主和配偶定位中的关键作用, 结果与荧光竞争结合试验高度一致, 进一步验证了分子对接技术的可靠性。同样, Guo等[25]利用分子对接技术, 证明了冷杉梢斑螟(Dioryctria Abietella)的气味结合蛋白DabiOBPs在摄食和产卵寄主定位中有重要作用, 展示了该技术在OBPs功能研究中的高效性。
在分子对接研究中, 结合能是衡量配体与受体结合能力的重要指标。研究表明, -4.5 kcal/mol通常被认为是具有生理活性的阈值, 结合能越小, 化合物具有生理活性的可能性越大, 但阈值附近的化合物也可能具有潜在活性[26]。分子对接技术的应用不仅仅局限于直观判断配体与受体的结合能力, 还能对结合能力强的挥发物进行分类, 实现大规模、快速地筛选与配体结合能力强的植物挥发物。同时, 通过分析挥发物的结构及官能团特性, 可改造结合能低的化合物以提升其与配体的结合能力。因此, 当从寄主中筛选大量候选挥发物, 并从中找出对靶标害虫嗅觉行为具有潜在调控能力的挥发物时, 分子对接技术可显著缩短传统实验周期、降低成本, 同时有助于解析蛋白质与挥发物之间的互作关系。
本研究选取蜂巢小甲虫触角中特异性高表达的OBP3为研究对象, 通过分子对接技术分析其与46种来自蜂巢材料和工蜂的挥发物的结合特性, 聚焦于5种已知影响蜂巢小甲虫行为的挥发物: (Z, Z, Z)-9, 12, 15-十八碳三烯酸乙基酯、十六烷酸乙酯、二十四烷、(Z)-9-十八碳烯酰胺和5-甲基-2-苯基-1H-吲哚。结果显示, 这5种挥发物与OBP3的结合能较低, 表明其与OBP3的结合能力较强, 可能在蜂巢小甲虫嗅觉反应中发挥重要作用, 介导其吸引或驱避效应。前人行为试验结果表明, 在合适浓度条件下, (Z, Z, Z)-9, 12, 15-十八碳三烯酸乙基酯和十六烷酸乙酯对蜂巢小甲虫有明显的引诱作用; 而二十四烷、(Z)-9-十八碳烯酰胺和5-甲基-2-苯基-1H-吲哚对蜂巢小甲虫表现出明显的驱避效应[19]。本研究的分子对接结果与这些行为学证据相呼应, 进一步支持OBP3在嗅觉信号识别中的潜在功能。
然而, 分子对接技术在嗅觉调控剂筛选中的局限性也不容忽视。本研究发现, 2-乙基己基-6-乙基-3-辛烷基邻苯二甲酸酯、3-甲基丁基十八烷基邻苯二甲酸酯、邻苯二甲酸, 壬基2, 4, 4-三甲基戊基酯等挥发物与OBP3结合能力很强, 可能也对蜂巢小甲虫具有类似嗅觉效应。但并没有在Dekebo等[19]的行为学试验中验证, 推测这可能与其研究方法有关, 该研究中使用了水蒸气蒸馏法提取化合物, 因此检测到一部分水溶性化合物以及常温下不挥发的物质。由于这几种物质尚未进行人工合成, 市面上无法获得, 因此本研究中未能开展行为学试验进行验证。这表明, 分子对接虽能预测结合能力, 但未充分考虑挥发物的理化特性(挥发性和稳定性)及行为学试验环境的复杂性。此外, 模型的准确性、配体选择偏差以及环境因素等也可能影响结果的可靠性。未来需构建“计算预测-物化筛选-行为验证”的闭环验证体系, 推动分子对接从定性筛选工具向定量预测平台的转变。
综上, 本研究通过同源建模和分子对接技术, 系统分析了蜂巢小甲虫触角特异性高表达OBP3与蜂巢材料及工蜂挥发物的结合特性, 结果表明OBP3可能在嗅觉识别和寄主定位中发挥重要作用, 为理解其生物学功能提供了重要依据。未来可结合RNAi技术和行为试验进一步明确OBP3具体作用机制, 为开发针对蜂巢小甲虫的嗅觉行为调控剂奠定基础。
-
本研究通过整合同源建模与分子对接技术, 阐明蜂巢小甲虫触角特异性高表达的OBP3蛋白对蜂巢材料及工蜂挥发物的广谱结合能力, 揭示该蛋白在害虫嗅觉介导的宿主定位中起核心作用。这一发现不仅从分子层面解析了OBP3的配体识别机制, 更为开发靶向嗅觉通路的害虫防控技术提供了关键理论支撑。