


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
多杀性巴氏杆菌(Pasteurella multocida,Pm)隶属巴氏杆菌科巴氏杆菌属,是一种两端钝圆,中间微凸,无芽孢,无鞭毛,有荚膜的败血性革兰阴性菌。Pm属条件性致病菌,正常情况下存在于动物口腔、消化道和呼吸道,一般不致病[1]。当天气骤变、饲养环境差(如通风不良、潮湿、拥挤、饲料霉变等)、营养缺乏、疲劳等应激因素出现导致动物免疫力降低时,该菌易突破机体的防御屏障,经淋巴液侵入黏膜并大量增殖,逃避宿主的固有免疫系统造成内源性感染[2]。该菌可导致动物呼吸系统疾病、局灶性感染及出血性败血症,如牛肺炎、禽霍乱、兔鼻瘘、有蹄动物的出血性败血症、牛羊猪等的船运热等[3]。该菌可引发猪肺疫,也可引起慢性感染造成严重的呼吸系统疾病(如猪萎缩性鼻炎),并诱发其他病原混合或继发感染,严重制约中国养猪业的健康发展[4-5]。
Pm致病力的强弱与其毒力因子有关。毒力因子是Pm在宿主体内存活、繁殖、致病的关键,有利于细菌入侵和定植,避免宿主启动防御机制,损伤细胞组织,刺激宿主产生炎症反应[1]。目前发现的毒力因子主要包括LPS、荚膜、OMPs、三聚体自转运蛋白、铁调节外膜蛋白(IROMPs)、脂蛋白E(PlpE)、皮肤坏死毒素(PMT)、脂蛋白B(PlpB)、Ⅳ型菌毛(PtfA)、丝状血球凝集素蛋白(FhaB)、黏附相关因子、蛋白酶等,根据其荚膜抗原和脂多糖,可分为A、B、D、E、F 5种血清群或1~16个血清型[6-7]。寡肽渗透酶A(OppA)是三聚体自转运蛋白家族的成员,在革兰阳性菌和阴性菌中均有发现,作为细胞质中的一种底物结合蛋白,其主要功能是捕获寡肽,或作为细胞膜的表面受体。常规培养条件(营养丰富的培养基)下,OppA属于低丰度蛋白,通常在菌体总蛋白中的占比小于1%,但当环境中寡肽匮乏时,Opp系统被激活,此时OppA的表达量显著上升,可达总蛋白的2%~5%。此外,该蛋白非常保守,与GenBank中收录的牛源、鹅源、鸡源等多株Pm进行比较,其相似性高于99%。文献[1]从福建部分猪场发病猪中分离出10株猪巴氏杆菌,其中6株为荚膜A型,4株为荚膜D型,并从分离株中成功克隆了OppA基因,相似性分析结果在97.27%~99.81%之间,进一步表明OppA基因具有很高的保守性,存在于国内报道流行的荚膜型中。OppA几乎存在于所有血清型Pm菌株中[8],但与其他种属细菌存在较大差异。因此,OppA可能是一个抵抗巴氏杆菌病的良好的交叉保护性抗原。目前OppA已在多种细菌研究中出现过报道,如:文献[9]对副猪嗜血杆菌的OppA基因进行了克隆、表达,并构建了间接ELISA检测方法;文献[10]对鼠伤寒沙门OppA的调控机理进行了分析;文献[11]对迟缓爱德华氏菌OppA蛋白进行了免疫原性研究;文献[12]证实了OppA是猪链球菌毒力因子之一。分析发现不同种属细菌OppA基因大小、蛋白结构均存在差异,且目前针对Pm OppA的研究鲜有报道,本试验通过对猪源Pm分离株的OppA基因进行克隆表达,对其免疫原性和免疫保护性进行初步探索,以期为开发相关防控制剂奠定前期基础。
全文HTML
-
猪源多杀性巴氏杆菌血清A型菌株(PM-1),分离自2022年9月重庆云阳某养殖场发病猪肺脏,由西南大学动物医学院动物疾病快速诊断中心鉴定并保存;SPF昆明鼠20只(20~25 g,雌性),购自重庆国家生物产业基地实验动物中心。
-
Easy Pure Viral DNA/RNA Ki(ER201-01)、FlyCut EcoR I(JE201-01)、FlyCut Xho I(JX201-01)和T4 DNA Ligase(FL101-01)均购自北京全式金生物技术股份有限公司;2×Phanta Flash Master Mix(Dye Plus)(P520-01)和DL2000 DNA marker(MD101-01)均购自南京诺唯赞生物科技股份有限公司;Prestained Protein Ladder(10-180 kDa)(AB116027)购自上海圣尔生物科技有限公司;Ni-NTA His Bind Resin(PANF001-005C)购自上海七海复泰生物科技有限公司;SanPrep核酸纯化套件(胶回收)(B51503)和SanPrep柱式质粒DNA抽提试剂盒(B518191)、BCA蛋白定量试剂盒(C503021)均购自生工生物工程(上海)股份有限公司;Top10感受态细胞、Rosetta感受态细胞和pET32a质粒保存于本实验室,山羊抗鼠IgG-HRP(SA00001-1)、小鼠IL-2 ELISA试剂盒(KE10100)和小鼠TNF-α ELISA试剂盒(KE10001)均购自Proteintech公司;弗氏佐剂(P2036/P2031)购自碧云天生物科技有限公司;小鼠抗巴氏杆菌阳性血清由西南大学动物医学院动物疾病快速诊断中心实验室制备。
-
凝胶成像仪(Viber Lourmat quantum-ST5)、蛋白成像系统(VILBER BIO IMAGING-Fusion FX5)、垂直电泳系统(CAVOY MP-8000)、多功能酶标仪(TECAN Infinite 200 PRO)、超声波细胞粉碎机(SCIENTZ-IID)、金属浴恒温加热器(Thermo Fisher)、高速冷冻离心机(6-10)、高速均质乳化仪(RCD-1A)、光学显微镜(CX31RTSF)。
-
利用
http://www.cbs.dtu.dk/services/TMHMM/ 在线工具中的Protparam分析OppA蛋白的基本理化性质;利用http://www.cbs.dtu.dk/serv-ices/SignalP/ 在线工具中的Signa1P 4.1 Server对亲疏水性进行预测;利用http://tools.immuneepitope.org/bcell/ 在线工具中的IEDB AnalysisResource预测B细胞表位。 -
参考GenBank中Pm UNMSM株OppA的基因序列(登录号为:KP660626.1),利用Primer Premier 5.0软件设计1对特异性引物。引物序列:OppA-F:5'-CCGGAATTCGCAGCAGAAGTACCC-3';OppA-R:5'-CCGCTCGAGTGTTCTTACCAGGATG3。扩增片段预期1 533 bp,斜体部分为EcoRⅠ和XhoⅠ酶切位点,引物由华大基因重庆分公司合成。利用一管式临床样品DNA抽提试剂盒提取纯化菌PM-1基因组,作为后续PCR反应的模板。基因OppA PCR反应体系:DNA模板4 μL,2×Taq PCR Master Mix 25 μL,ddH2O 13 μL,上、下游引物(10 μmol/μL)各4 μL。PCR扩增条件为:95 ℃预变性3 min,95 ℃变性15 s,55 ℃退火15 s,72 ℃延伸15 s,37个循环,72 ℃终延伸5 min。PCR产物以1%凝胶电泳并利用胶回收试剂盒纯化,阳性产物送至华大基因重庆分公司测序。
-
利用限制性核酸内切酶FlyCut EcoRⅠ和FlyCut XhoⅠ分别对PCR扩增产物和pET-32a载体进行酶切反应,胶回收黏端片段,利用T4 DNA连接酶将OppA片段与pET-32a载体进行连接,并转化至TOP10感受态细胞中,涂布LB固体培养基(含1% Amp),37 ℃过夜培养,挑取单菌落进行PCR鉴定,阳性菌利用质粒提取试剂盒提取重组质粒,测序鉴定正确后命名为pET32a-OppA,保存于-20 ℃备用。
-
将质粒pET32a-OppA转化至BL-21感受态细胞,阳性重组子接种于LB液体培养基(含1% Amp)中恒温振荡培养4 h左右,测定菌液OD600 nm值,待至0.6~0.8后加入1%比例IPTG (1 m mol/L),继续180转37 ℃振荡培养4 h左右。取出置于4 ℃终止诱导。将菌液置于4 ℃,以8 000r/min的速度离心5 min,然后收集沉淀和上清液。使用PBS将菌体沉淀充分重悬后,使用细胞破碎仪破碎3~5次,破碎产物于4 ℃,8 000r/min离心收集沉淀。沉淀中加入20 mL预冷的快速洗涤液充分吹打混匀,静置10 min后于4 ℃,8 000r/min离心收集沉淀,再加入20 mL预冷的快速溶解液使蛋白变性,最后取10 mL表达产物进行透析复性。
-
将诱导型表达的重组蛋白通过Ni-NTA重力柱进行蛋白纯化,纯化后的洗脱液进行SDS-PAGE电泳,再经考马斯亮蓝染色后鉴定结果。纯化蛋白进一步通过Western blot验证其生物活性,经SDS-PAGE电泳后转印至NC膜,以小鼠抗巴氏杆菌阳性血清作为一抗,HPR标记的山羊抗鼠IgG作为二抗进行Western blot,利用DAB显色试剂盒显色并记录结果。
-
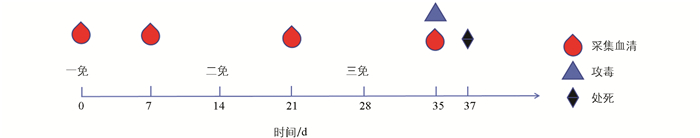
动物试验方案如图 1所示,20只雌性SPF小鼠随机分为2组,每组10只,分别为免疫组和未免疫组。其中免疫组注射弗氏佐剂乳化的OppA蛋白,分别于第0 d、14 d、28 d通过背部皮下注射进行3次免疫,免疫剂量均为12 μg/只;未免疫组均注射等量无菌生理盐水。两组小鼠在第0 d(免疫前)、7 d,21 d和35 d共4个时间点采集尾静脉血,分离血清,通过间接ELISA方法检测其OppA特异性抗体水平,并利用试剂盒对细胞因子IL-2和TNF-α的含量进行检测。OppA抗体间接ELISA检测方法步骤如下:利用BCA蛋白浓度测定试剂盒测定OppA蛋白浓度,按照1 μg/mL浓度包被至96孔酶标板,4 ℃过夜;利用PBST洗涤5次,每次1 min;拍干后每孔加入100 μL 5%脱脂奶粉置37 ℃封闭2 h,甩干孔内液体,利用PBST洗涤5次,每次1 min;拍干后加入20倍稀释的待检小鼠血清100 μL,37 ℃孵育30 min,重复洗涤后加入1∶6 000稀释的HRP羊抗鼠IgG(酶标二抗),每孔100 μL,37 ℃孵育30 min,重复洗涤;每孔加入100 μL TMB显色液,避光37 ℃孵育15 min,最后每孔加入50 μL终止液,酶标仪测定各孔的OD450值。细胞因子IL-2和TNF-α的检测按照试剂盒说明书进行。
-
攻毒之前,挑取PM-1菌落在液体培养基中培养,测量菌液OD600nm值,待其约0.8时取出。梯度稀释PM-1菌液并涂布平板过夜培养,观察平板并选择合适稀释倍数的分离菌悬液进行攻毒。预试验结果显示小鼠感染剂量为1×106 CFU/只时小鼠不会迅速死亡并表现明显临床症状,便于观察攻毒后小鼠临床表现并记录存活时间。第35 d采血后,两组小鼠腹腔感染PM-1菌,感染剂量为0.2 mL(1×106 CFU/只)。分离菌悬液,分别记为免疫攻毒组与未免疫攻毒组。感染后每4 h观察一次小鼠的临床表现,共连续观察48 h,统计各组小鼠的存活时间,绘制存活曲线图。小鼠死亡后立即进行解剖,取肝、脾、肺3种脏器进行病理组织学观察,具体步骤参考文献[13]进行,组织切片经HE染色后置显微镜下观察病变。
-
试验数据均利用GraphPad Prism9.0软件进行统计学分析及作图。采用方差分析(ANOVA)比较免疫组与空白组间的差异,p<0.05为差异显著。小鼠攻毒后存活曲线采用Log rank检验分析,p<0.05为差异显著。
1.1. 菌株、实验动物来源
1.2. 主要试剂
1.3. 主要仪器
1.4. OppA蛋白的生物信息学分析
1.5. 猪源多杀性巴氏杆菌OppA基因克隆
1.6. OppA基因重组质粒的构建与鉴定
1.7. OppA基因的诱导型表达及变复性
1.8. OppA重组蛋白鉴定及免疫活性的验证
1.9. OppA重组蛋白免疫小鼠血清中IgG抗体和细胞因子的检测
1.10. OppA重组蛋白对小鼠的免疫保护性试验
1.11. 数据处理与统计学分析
-
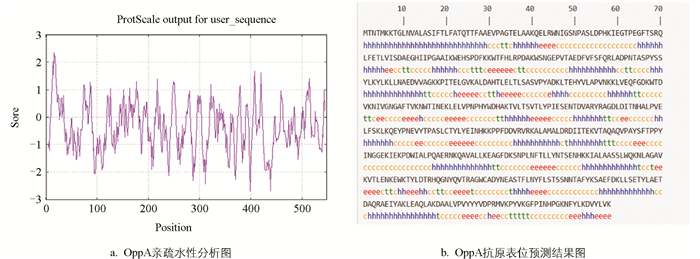
分析显示OppA蛋白由548个氨基酸组成,其中占比最高的蛋白是丙氨酸(10.6%),脂溶指数83.72,亲水性总平均值-0.368,不稳定性指数为29.48,判定该蛋白为稳定蛋白;亲疏水性预测显示,OppA蛋白高疏水性区域位于第16位氨基酸附近,高亲水性区域位于第399位氨基酸附近(图 2a)。抗原表位预测显示,OppA蛋白有B细胞抗原表位,其中优势表位为第11~21、72~78、119~127位等21个抗原表位(图 2b)。
-
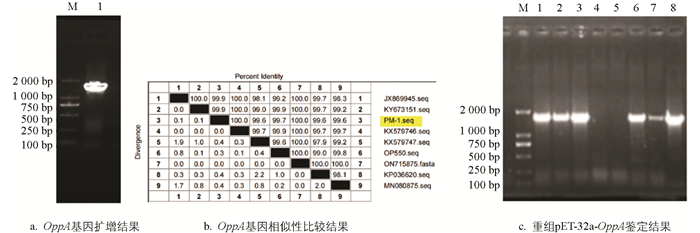
PCR扩增得到大小1 533 bp的目的条带(图 3a),与预期结果一致;测序后利用NCBI数据库中BLAST功能进行相似性比对分析,结果显示猪源Pm OppA基因与Genbank中收录的牛源(ON715874)、山羊源(ON715875)、鸭源(JX869945、KX579747)、鸡源(KY673151)、鹅源(KX579746、KP036620)和人源(MN080875)的Pm相似性在99.6% ~100%之间(图 3b),说明OppA基因克隆成功。将OppA基因与pET32a载体连接,转化后挑取单克隆进行PCR鉴定,结果如图 3c所示,显示8个克隆中有6个为阳性,阳性重组子经测序后显示构建成功,菌株于-20 ℃保存备用。
-
IPTG诱导猪源OppA蛋白表达纯化后进行SDS-PAGE电泳检测,结果(图 4a)显示,OppA重组蛋白大小约为74 kDa,与预期大小一致,且经过纯化,在其洗脱液中杂蛋白条带明显减少。Western blot鉴定结果(图 4b)显示,OppA重组蛋白能够与小鼠抗巴氏杆菌阳性血清发生反应,说明该蛋白具有免疫活性。
-
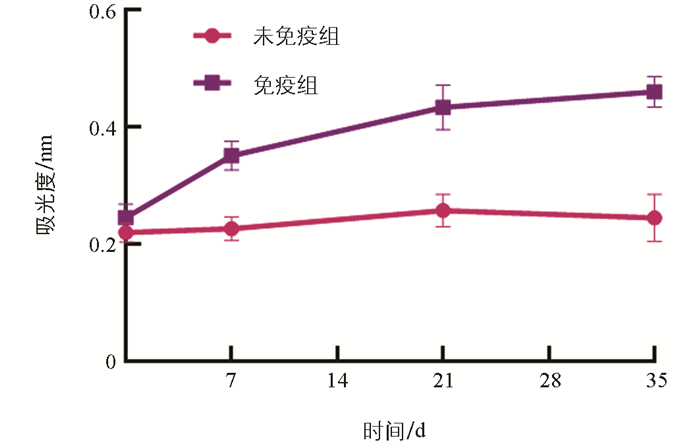
以复性后OppA作为包被抗原,利用间接ELISA方法对0 d、7 d、21 d和35 d小鼠血清中OppA特异性抗体水平进行检测。测定结果(图 5)显示,一免后第7 d,免疫组小鼠血清中抗体水平显著上升,与空白组小鼠差异显著(p<0.05);二免、三免过后,免疫组小鼠血清中抗体水平进一步升高,均显著高于空白组(p<0.05)。未接种免疫组小鼠血清中相应抗体水平自始至终变化不明显。
-
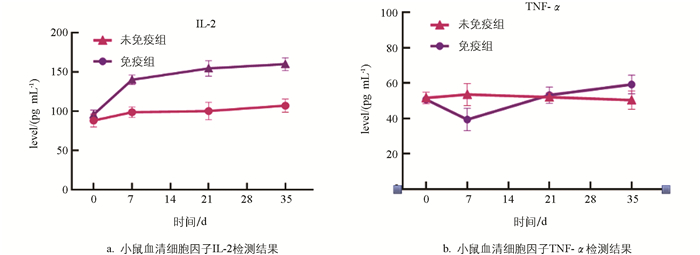
利用ELISA试剂盒对小鼠血清中细胞因子IL-2和TNF-α的含量进行检测,结果发现免疫组小鼠IL-2在一免后迅速上升(图 6a),二免、三免后仍现缓慢上升趋势,第35 d时仍保持较高水平,在第7 d、21 d和35 d 3个时间点均显著高于未免疫组小鼠(p<0.05);而未免疫组小鼠血清中IL-2含量均无显著变化。血清中TNF-α检测结果如图 6b所示,一免后第7 d其含量显著低于未免疫组(p<0.05),但在第21 d和35 d时其含量又逐渐上升,与未免疫组之间差异不显著(p>0.05)。
-
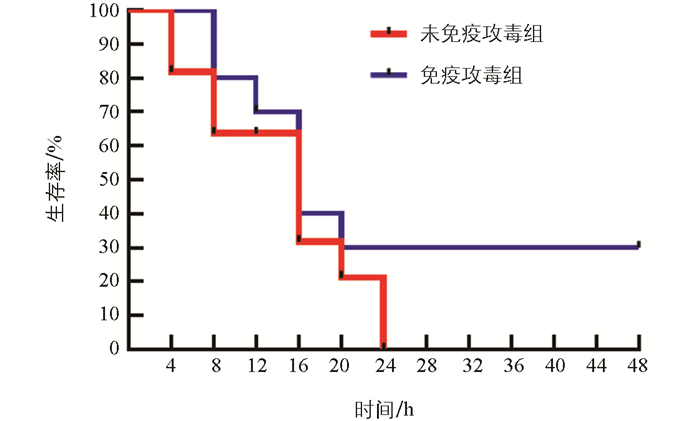
第35 d,两组小鼠攻毒后均出现精神萎靡,双眼微闭,被毛凌乱的现象。未免疫攻毒组小鼠最先(攻毒后4 h)出现死亡,并在24 h内100%死亡(图 7);免疫攻毒组小鼠在攻毒后8 h出现死亡,24 h内共死亡7只,其余3只小鼠在观察期内均存活,该组死亡率为70%,尽管存活率高于未免疫接种组,但统计学分析显示两组之间差异不显著(p>0.05)。
-
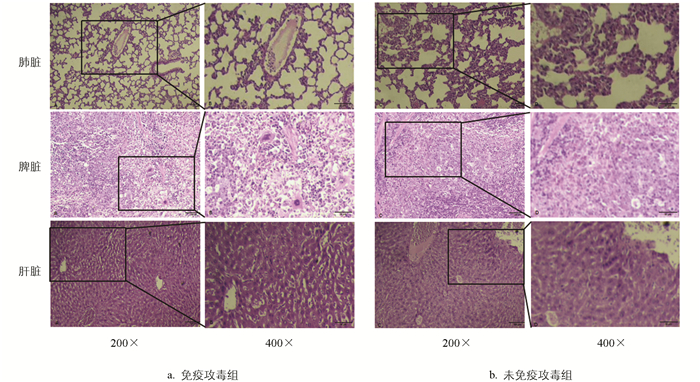
攻毒后死亡小鼠肺、脾、肝脏等组织病理变化如图 8所示。未免疫攻毒组小鼠肺脏组织中发现肺泡上皮增生,肺泡壁变厚,肺泡内有大量炎性细胞浸润,并伴有明显的出血现象;脾脏组织出血,炎性细胞浸润;肝脏组织明显出血,血管扩张,血细胞溢出,肝细胞肿胀。免疫攻毒组小鼠的病理特征与未免疫攻毒组相似,但病变程度相对较轻,如肺脏、脾脏有轻微出血和炎性细胞浸润,肝脏出血、水肿等。
2.1. OppA基因生物学软件分析
2.2. 猪源Pm OppA基因克隆及原核表达载体的构建
2.3. 猪源Pm OppA重组蛋白表达与免疫活性鉴定
2.4. 小鼠血清OppA特异性抗体水平检测结果
2.5. 小鼠血清中细胞因子检测结果
2.6. 攻毒后小鼠存活时间
2.7. 攻毒小鼠组织病理学观察
-
多杀性巴氏杆菌的血清型较多,感染宿主谱非常广泛,目前在我国感染猪的血清型主要是A和D型[14],国内也有商品化的疫苗,主要以灭活苗和弱毒苗为主。然而灭活疫苗对不同血清型的多杀性巴氏杆菌的交叉免疫保护力较差,需要较大的免疫剂量才能达到可靠的保护率;弱毒苗存在毒力返祖返强的风险,在储存和长途运输中存在着一定的弊端,并且与我国现阶段流行的优势血清型不符,导致免疫效果并不理想[15]。鉴于此,开发新型防控制剂尤为重要,而重组亚单位疫苗因不含病原体的“活”成分,相较于含有完整病原体的疫苗更安全和稳定,且可针对多种血清型,因此开展猪多杀性巴氏杆菌的亚单位疫苗的研制非常重要。
通常一些细菌组分(尤其是分泌蛋白和外膜蛋白)和致病菌的毒力因子被认为是亚单位疫苗的候选靶标[16-17],而OppA是多杀性巴氏杆菌中重要的跨膜蛋白,在不同血清型菌株中均存在,且在同种细菌中高度保守,业已证实在鼠伤寒沙门菌[10]、迟缓爱德华菌[11]、副猪嗜血杆菌[18-19]中能引起特异性免疫应答,对宿主具有一定的保护作用。本试验首先通过生物信息学软件对多杀性巴氏杆菌的OppA蛋白的亲水性及细胞表位进行分析,推测其也应该具有较强的免疫原性,进而通过原核表达系统将该蛋白进行表达,复性后的蛋白经Western blot鉴定能够与小鼠抗巴氏杆菌阳性血清发生结合反应,说明该蛋白具有生物学活性。
抗体是机体抵抗多杀性巴氏杆菌感染和产生免疫保护的主力,抗体水平是验证其抗原蛋白免疫原性及其诱导机体产生体液免疫应答的主要指标[20-21]。本试验采用间接ELISA方法对3次免疫后小鼠血清中OppA特异性抗体进行测定,发现在首免后7 d即可产生高滴度的抗体,两次加强免疫均能使其抗体维持在较高水平,说明OppA能够诱导小鼠产生强烈的体液免疫应答反应。IL-2和TNF-α均属于Th1类细胞因子,研究表明IL-2是引起T细胞增殖的主要细胞因子,它具有促进T、B淋巴细胞的增殖和分化,增强单核细胞以及NK细胞的杀伤活性等免疫调节功能,主要介导细胞免疫应答,调节细胞毒性T细胞分化和参与迟发型超敏反应[22]。TNF-α是重要的炎性因子,主要由单核巨噬细胞产生,当其与受体结合后启动细胞内信号,诱导激活炎性细胞聚集,导致细胞坏死和凋亡而引起急性炎症反应[23]。本试验对3次免疫OppA蛋白小鼠血清中的IL-2和TNF-α进行检测,发现一免后IL-2的含量迅速上升,二免、三免后仍呈现缓慢上升趋势,35 d时仍保持较高水平,究其原因,其一是OppA蛋白能够活化机体T细胞的增殖,促进其分化,能够诱导宿主的细胞免疫,其二可能是制作抗原不纯,残留LPS导致IL-2含量迅速上升。炎性因子TNF-α在一免后显著下降,二免和三免后与对照组差异不显著,说明多杀性巴氏杆菌OppA蛋白不会引起机体的炎性反应。这一结果与文献[19]利用副猪嗜血杆菌重组OppA蛋白进行小鼠免疫保护性试验结果一致。
保护性试验显示OppA蛋白免疫组小鼠的存活率为30%,未免疫组全部死亡,说明OppA蛋白虽对小鼠有一定的保护性,但保护率较低。进一步通过观察死亡小鼠主要脏器的病理组织学变化,发现无论是否免疫OppA蛋白,其病理变化均十分明显,尽管免疫组的病变略显轻微,但均已造成脏器的损伤,这一结果与文献[24]利用副猪嗜血杆菌重组OppA蛋白对猪进行保护性试验的结果十分相似,文献[11]利用OppA刺激大菱鲆产生免疫应答,尽管延长了免疫组死亡时间,但最终的免疫保护率为25.9%。究其原因,其一是因为细菌的毒力因子众多,仅凭单个蛋白尽管能够激发机体的免疫系统,但仍不能产生足够的保护力;其二,实验动物小鼠对多杀性巴氏杆菌极为敏感,当免疫系统产生的细胞免疫和体液免疫不足以清除所有的细菌时,残余的多杀性巴氏杆菌足以对其产生致死性后果。因而本试验结果证实OppA蛋白对小鼠具有一定保护性,但若将其开发为亚单位疫苗,可能需要联合多个保护性抗原才能取得更好的保护作用。目前已有对猪多杀性巴氏杆菌OmpW、TbpA蛋白的免疫原性进行的研究报道,发现OmpW和TbpA蛋白在不同血清群猪多杀巴氏杆菌中均高度保守,且攻毒保护试验表明OmpW和TbpA蛋白对小鼠的保护率分别为50.0%和37.5%[25]。OppA蛋白可与不同抗原蛋白进行组合,筛选最佳组合方案,以获得免疫效力较强的亚单位疫苗。
综上所述,本试验对猪源多杀性巴氏杆菌OppA蛋白进行了原核表达及免疫原性的初步探索,结果显示该蛋白具有良好的免疫原性,能够激发小鼠产生免疫应答反应,具有一定的免疫保护效果。