


 下载:
下载:


-
开放科学(资源服务)标识码(OSID):

-
苹果是世界四大水果之一,其产业健康发展对全球果品供应至关重要。盐胁迫会引发离子毒害、渗透胁迫和氧化胁迫,严重抑制苹果生长发育进而影响苹果的产量和品质[1]。因此,解析苹果的耐盐机制并开展盐胁迫条件下的苹果育种研究,对推动相关产业发展具有重要的现实意义。目前,关于苹果的研究多集中于形态建成与激素调控,而对其抗逆性,特别是耐盐机制的研究则相对薄弱。
油菜素甾醇(Brassinosteroids,BR)是调控植物生长发育和逆境响应的关键激素,其信号通路的核心转录因子BZR1在介导非生物胁迫响应中扮演着重要角色。研究表明:在盐胁迫下,BZR1可通过调控活性氧清除系统,如超氧化物歧化酶(Superoxide Dismutase,SOD)等相关基因的表达来增强植物的氧化耐受性,这是植物耐盐性的关键机制之一[2-5]。油菜素内酯(Brassinolide,BL)是BR家族中首个被分离鉴定的成员,因其具有最高的生物学活性,且其生理功能与分子信号通路表征最为明确,故而被视为该类植物激素的典型代表。外源施用活性BR类似物,如2,4-表油菜素内酯(2,4-Epibrassinolide,EBR)能显著增强多种植物对逆境胁迫的耐受性[6-8]。植物遭遇非生物胁迫时,通过外施EBR能够显著减轻其受到的负面影响。在黄瓜(Cucumis sativus L.)、水稻(Oryza sativa L.)和玉米(Zea mays L.)等作物上的研究表明:EBR增强逆境耐受性的机制依赖于其对植株H2O2的清除作用[9-10],涉及NO(Nitric Oxide)的调控[11]。同时,EBR处理可有效促进植物渗透调节能力,显著激活SOD、过氧化物酶(Peroxidase,POD)等抗氧化酶系统的活性,提高光合作用效率[12-14]。这些协同作用的生理响应共同构成了植物应对非生物胁迫的综合防御机制,从而大幅提升其在逆境条件下的生存能力[15-16]。
BR功能的发挥依赖于其内源水平的精确调控。在BR生物合成途径中,DWF4基因编码的C-22羟化酶催化限速步骤,是控制BR内源水平的关键枢纽。研究表明:DWF4不仅调控细胞伸长、分裂和生物量积累,更在非生物胁迫应答中起重要作用[17-19]。然而,上述功能研究多集中于模式植物和草本作物。以苹果为代表的木本植物中,具有特殊株型的苹果材料尤为典型,其MdDWF4基因如何通过差异性表达调控,MdBZR1转录因子是否通过直接结合并调控MdDWF4基因的启动子来协同增强耐盐性,均属未知。因此,研究以柱型苹果为材料,聚焦BR合成关键基因MdDWF4,旨在解析其表达模式、外源EBR的生理效应及生长调控功能,最终阐明MdDWF4通过动态调控BR水平介导耐盐性的分子机制,为苹果抗逆遗传改良提供新靶点与理论依据。
全文HTML
-
试验中植物材料:柱型苹果品种“威赛克旭(Wijcik McIntosh)”(Malus domestica),以及野生型拟南芥品种“哥伦比亚(Col)”(Arabidopsis thaliana)。
拟南芥在环境温度23 ℃、湿度70%、光照时长16 h中培养,盐胁迫处理采用300 mmol/L NaCl溶液。观察胁迫0、4、7、14、40和50 d的拟南芥植株状态,并取拟南芥叶片,测定相关生理指标。试验所得数据均通过3次独立的生物学重复获得。
-
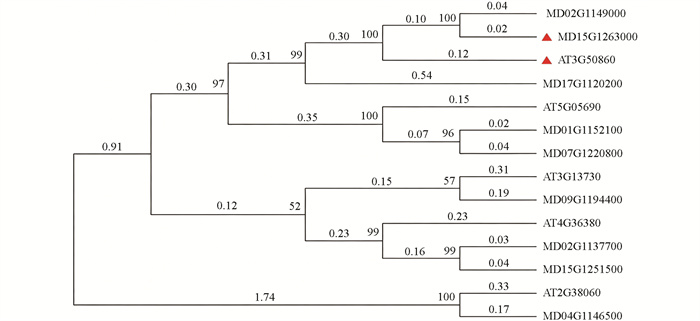
利用MEGAX7.0软件,采用邻接法(Neighbour Joining,NJ)对拟南芥、胡杨、水稻等多个物种已发表的DWF4基因进行氨基酸序列比对,并构建系统进化树。
-
分别从柱型苹果叶片和转基因拟南芥的整株样本中提取RNA。采用实时荧光定量技术,在热循环仪(CFX Connect Real-Time System;Bio-Rad,Hercules,CA)上进行qRT-PCR检测。基因表达定量分析使用Bio-Rad CFX96 Touch实时荧光定量PCR仪,以内源性参照基因MdActin作为内参。反应体系为:2x Pre STAR MAX 10 μL,上下游引物(10 μmol/L)各1 μL(表 1),cDNA 4 μL,用dd H2O补足至20 μL。扩增程序为:94 ℃预变性3 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸90 s,循环数为34;最后72 ℃延伸5 min[20-21]。通过2-ΔΔCT法定量基因表达水平,所有数据源自3次独立重复试验并计算相对表达量及其标准差(SD)。试验数据通过SPSS软件(IBM SPSS Statistics 25,USA)进行t检验和ANOVA单因素方差分析(p<0.05)。
-
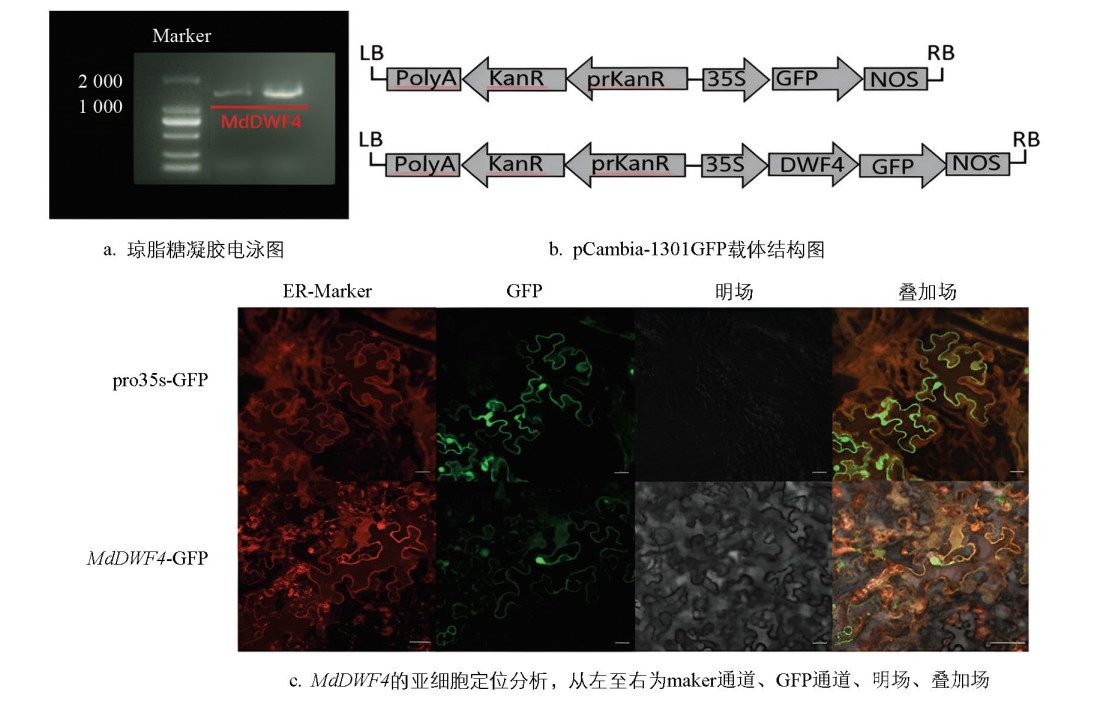
克隆去除终止密码子的MdDWF4基因,并将其连接至绿色荧光蛋白表达载体的N端,成功构建MdDWF4 -GFP融合表达载体。通过含利福平、庆大霉素和卡那霉素抗性的LB培养基筛选阳性克隆,经PCR鉴定正确后,用含10 mmol/L MES、10 mmol/L MgCl2、100 μmol/L AS液体培养基重悬农杆菌至OD600约0.6,通过瞬时转化法[11-12]将pCambia-1301GFP载体注射到本氏烟草叶片中观察。空载体与MdDWF4基因载体均表现出正常的荧光信号。
-
试验中使用的高保真酶PCR Mix为TAKARA公司生产的primer STAR产品(货号:R045A)。以含MdDWF4的克隆质粒为模板,亚细胞定位引物PCR扩增(表 2)。载体pCambia-1301GFP经过TAKARA公司提供的Bam HI限制性内切酶,单酶切处理,酶切产物依次进行胶回收纯化、连接转化、阳性克隆筛选及测序验证,最终构建重组载体。选取25龄的本氏烟草植株,在光照培养室中种植,进行瞬时转化法注射后标记叶片,经24 h暗培养和24 h光培养后,取下表皮层的烟草叶片,用激光共聚焦显微镜进行观察。
-
载体选用pCambia-1301,限制性内切酶为PstI,引物序列见表 2。野生型拟南芥植株去顶并生长3 d后,使用农杆菌(OD600=1.2~1.6)进行侵染。转化后收集的T0代种子经过表面灭菌,均匀播种在1/2 MS选择培养基上。培育3代获得过表达的纯合株系,从6个株系中各取3株整株样品提取RNA,使用引物QMdDWF4-F/QMdDWF4-R进行荧光定量PCR,内参引物为AtUBQ-F/AtUBQ-R。
-
将PHIS- MdDWF4 (含启动子)与ADT7- MdBZR1 (含诱饵蛋白)共转化至酵母Y187感受态细胞,转化采用PEG/LiAc法,经过热激处理后,将细胞涂布于SD/-Leu/-Trp选择性培养基上筛选共转化阳性菌落。阳性菌落转接至含3-氨基三唑(3-AT,80 mmol/L)的SD/-His/-Leu/-Trp选择性培养基,以抑制背景自激活。观察SD/-His/-Leu/-Trp选择性培养基上菌落的生长情况。
1.1. 材料
1.2. 系统进化树
1.3. 表达分析
1.4. MdDWF4蛋白的亚细胞定位
1.5. MdDWF4基因的基因克隆与载体构建
1.6. MdDWF4基因的过表达载体构建与拟南芥遗传转化和鉴定
1.7. 酵母单杂交(Y1H)
-
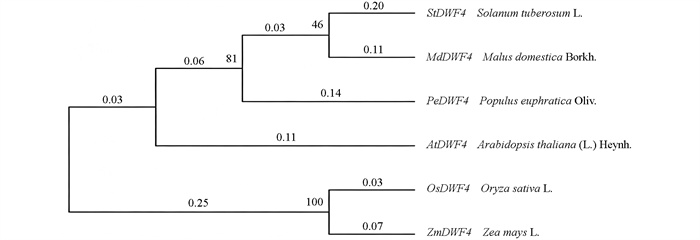
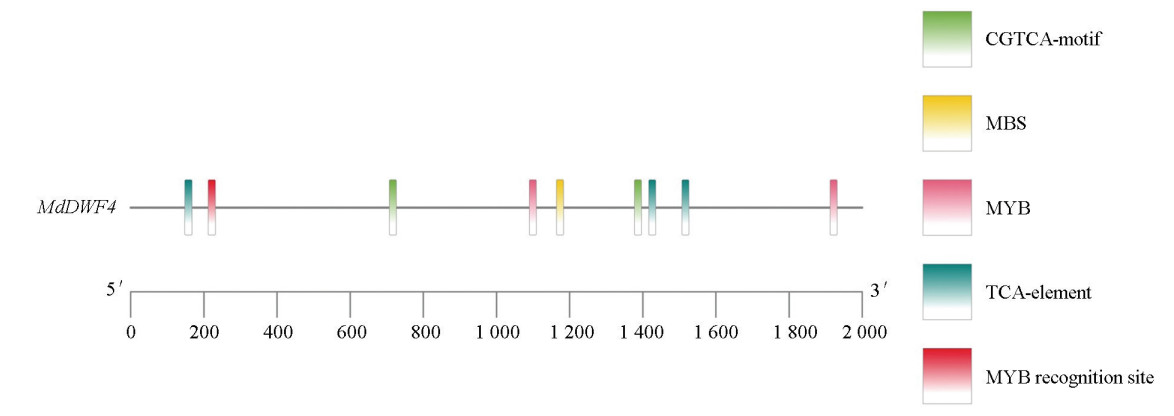
通过同源比对与系统进化分析,从柱型苹果中鉴定出一个与拟南芥AtDWF4亲缘关系最近的基因,并将其命名为MdDWF4 (图 1)。序列分析表明,MdDWF4基因的编码区(CDS)全长为1 482 bp,编码一个由493个氨基酸组成的蛋白质。对该蛋白进行保守结构域预测发现,其包含典型的细胞色素P450超家族结构域(图 2)。进一步探究其进化保守性,将其与拟南芥、土豆、水稻等多个物种的DWF4蛋白进行了多重序列比对。结果显示,这些蛋白之间存在多个短且高度保守的氨基酸片段,且这些序列中以高度一致的线性顺序排列,通过系统进化树分析进一步支持了它们功能的相似性(图 3)。上述结果表明:DWF4基因在进化过程中高度保守。此外,对MdDWF4基因启动子序列(起始密码子ATG上游约2 000 bp)的分析发现,其含有多个潜在的顺式作用元件,包括2个参与茉莉酸甲酯应答的CGTCA-motif、1个与非生物胁迫相关的MBS元件,以及其他与水杨酸、脱落酸等激素应答相关的元件(图 4)。研究结果表明:MdDWF4的表达可能受到多种激素与非生物胁迫的复杂调控。
-
通过荧光信号叠加分析,绿色与红色荧光在相同位置完全重合,进一步证明了MdDWF4蛋白定位于内质网(图 5)。这一结果与真核细胞色素P450酶通常定位于内质网的研究结果一致。由于MdDWF4催化BL合成途径中的限速步骤,其在内质网的定位与植物细胞内质网合成BR的假设高度吻合,进一步支持了MdDWF4在BR合成中的关键作用。
-
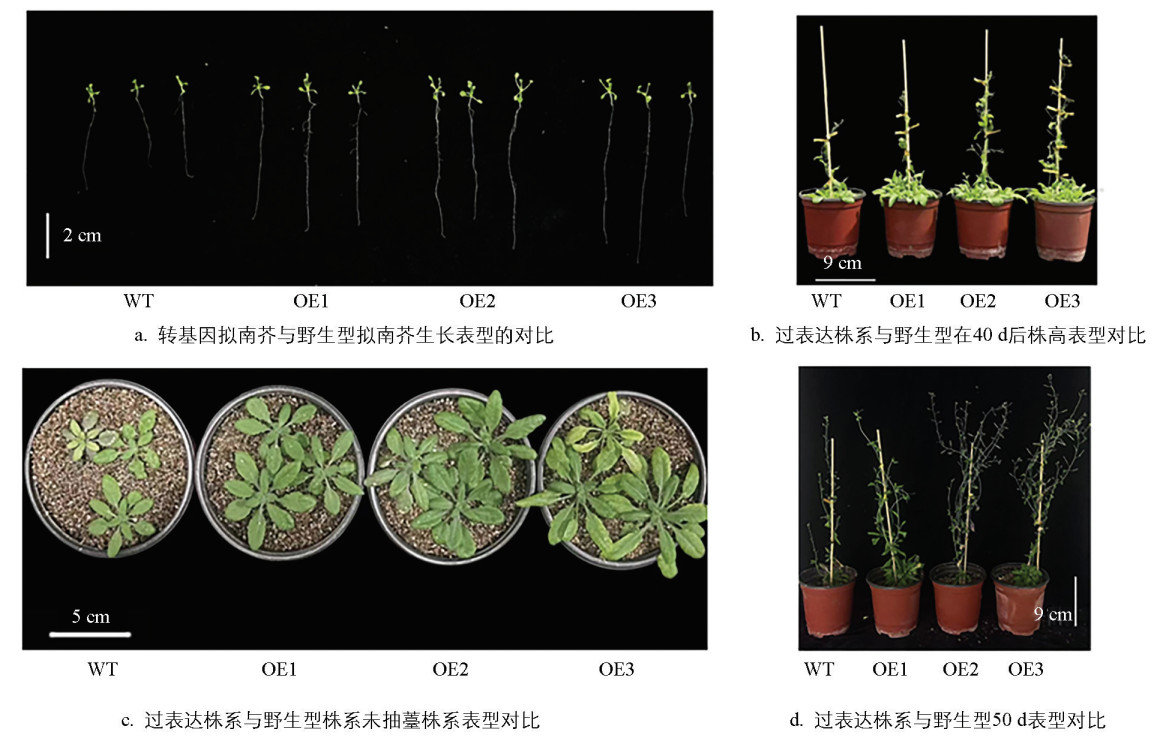
通过对过表达MdDWF4的转基因拟南芥的表型观察分析,表明野生型与转基因拟南芥的根长在14 d后的表现存在显著差异。野生型拟南芥的平均根长为35.5 mm,而转基因拟南芥分别为54.3、57.8、59.1 mm,分别显著增加了52.96%、62.82%、66.48%。继续培养14 d后,过表达株系和野生型叶片形状相同,但是生物量显著增加。野生型植株鲜质量为32.6 mg,3个过表达株系OE1、OE2、OE3鲜质量平均分别为45.3、51.6、68.0 mg,与野生型相比分别增加了38.95%、58.28%、108.58%(图 6)。
在试验进行至第40 d时,观察到拟南芥进入生殖生长阶段,顶端开始开花并形成果荚(图 6b)。对野生型和不同过表达株系的株高进行测量结果显示,野生型植株的平均高度为11.26 cm,而过表达株系的株高分别为14.31、17.00、22.96 cm,较野生型拟南芥显著增加了27.08%、50.97%、60.44%。进一步分析表明:过表达株系的表型变异程度与MdDWF4基因的表达水平呈显著正相关,证明了MdDWF4基因的过量表达是拟南芥表型改变的主要因素。到第50 d时,过表达植株的果荚大部分已成熟,而野生型拟南芥果荚则多数未成熟,且过表达拟南芥的分枝数明显多于野生型拟南芥(图 6d)。
-
为验证MdDWF4在植物盐胁迫响应中的功能,对野生型拟南芥及其MdDWF4过表达株系OE1、OE2、OE3进行了盐胁迫处理。在300 mmol/L NaCl溶液处理4 d后,野生型植株出现典型的盐胁迫症状,包括叶缘脱水、叶片黄化及生长停滞等现象。相比之下,过表达株系的萎蔫程度明显减轻,OE1株系(其MdDWF4表达量低于OE2和OE3)表现出稍优于野生型的生长状态,但差异不显著;而OE2和OE3株系则表现出显著的抗盐性,叶片保持较好的水分和绿色程度。上述结果初步证明MdDWF4能增强拟南芥的盐胁迫耐受性。
-
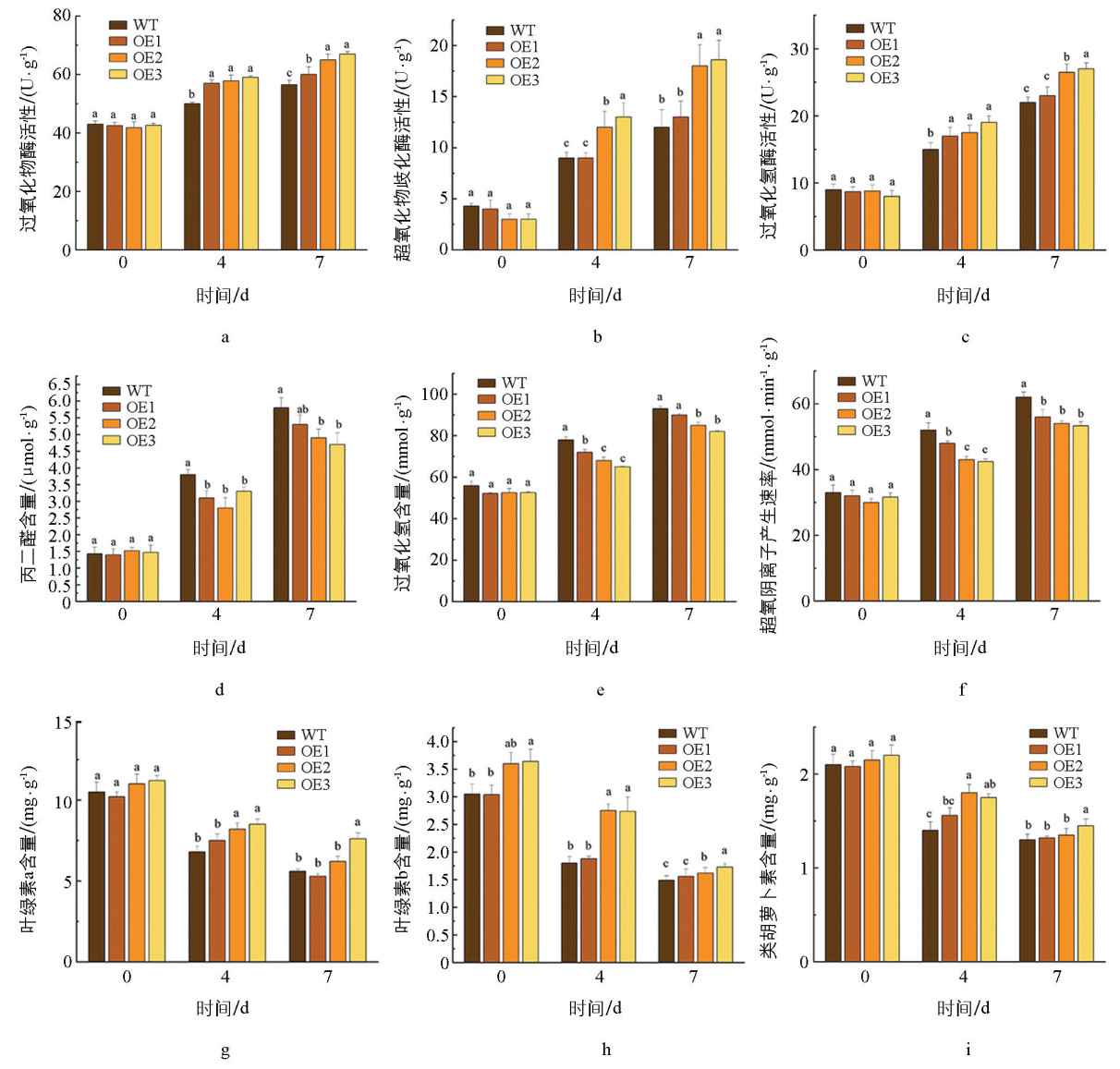
盐胁迫诱导的活性氧(ROS)暴发是造成细胞损伤的关键因素。随着盐胁迫时间的延长,野生型植株的过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)虽有一定的升高,但其清除能力不足,导致超氧阴离子(O2-)产生速率和过氧化氢(H2O2)含量急剧上升。过表达株系则展现出更强的抗氧化潜力:其POD、CAT和SOD的活性较野生型显著升高(图 7a、b、c),同时O2-的产生速率和H2O2含量增幅较野生型显著下降。作为细胞膜脂质过氧化的重要标志物,丙二醛(Malondialdehyde,MDA)含量在过表达株系中显著低于野生型,进一步证明了MdDWF4在缓解膜脂过氧化中的作用(图 7d、e、f)。这些结果表明:MdDWF4能正向调控抗氧化酶的活性,增强植物对ROS的清除能力,从而缓解植株在盐胁迫下受到的氧化损伤和膜脂过氧化。
-
氧化损伤的减轻有助于保护光合器官。测定光合色素,发现盐胁迫条件下,拟南芥叶绿素a(ChlorophyllA,Chla)、叶绿素b(ChlorophyllB,Chlb)和类胡萝卜素(Carotenoid)的含量显著降低,说明盐胁迫对光合系统产生了严重的破坏。与此相对,过表达株系的3种关键合成色素含量明显高于野生型植株,这表明MdDWF4的过表达有助于减轻盐胁迫对光合作用系统的损害(图 7g、h、i),直接保护了光合机构,维持了光合色素的稳态,这为转基因植株在盐胁迫下能保持更好的光合能力与绿色表型提供了生理基础。
-
鉴于BZR1在整合盐胁迫等信号中的核心作用,推测MdBZR1可能直接调控MdDWF4的转录。酵母杂交试验结果显示,在不同培养基条件下观察到明显的生长差异(图 8)。在SD/-Leu/-Trp选择性培养基中,试验组、阳性对照和阴性对照均显示酵母生长;而在SD/-His/-Leu/-Trp选择性培养基中,试验组和阳性对照表现出生长活性,阴性对照则未表现出生长。这一现象表明:MdBZR1转录因子能够特异性结合并调控MdDWF4基因启动子区域,进而影响其表达。
2.1. 苹果MdDWF4基因的生物信息学分析
2.2. MdDWF4蛋白的亚细胞定位
2.3. 过表达MdDWF4基因的拟南芥的表型分析及鉴定
2.4. 过表达MdDWF4提升拟南芥的盐胁迫抗性
2.4.1. 过表达MdDWF4激活拟南芥抗氧化系统缓解氧化损伤
2.4.2. 过表达MdDWF4缓解拟南芥盐胁迫对光合系统的损伤
2.5. MdBZR1与MdDWF4启动子互作
-
本研究结果表明:MdDWF4作为油菜素内酯生物合成的限速酶,是柱型苹果中协同调控正常生长与盐胁迫耐受性的关键因子。这一结论得到了转基因拟南芥试验的支持,过表达MdDWF4不仅能重现典型的BR过量表型(如株高、生物量显著增加),更显著增强了植株的耐盐能力,提示MdDWF4在生长与抗逆协同调控中具有普适功能。
通过对MdDWF4的克隆与生物信息学分析结果显示:该基因编码区的序列长度为1 482 bp,其编码蛋白质由493个氨基酸组成。亚细胞定位结果证明了MdDWF4蛋白定位于内质网。分析系统进化树发现其与马铃薯、胡杨等物种的DWF4同源蛋白高度相似[22],因此推测DWF4在植物中可能具有相似的、保守的生物学功能。此外,本研究不仅通过生物信息学分析佐证了不同物种间DWF4蛋白功能的保守性,也通过转基因手段验证了MdDWF4的生物学功能。在拟南芥中异源过表达MdDWF4所引发的促生长表型,与前述物种中的报道一致[23-24]。MdDWF4蛋白功能在玉米、水稻、毛白杨和胡杨等物种中高度保守。不同物种的DWF4同源基因在调控植物生长发育、非生物胁迫响应及光合作用中具有保守功能,体现了DWF4蛋白结构域的保守性。同时,BR生物合成基因(如DWF4)可显著增强植物对多种胁迫的耐受性[25]。更为重要的是,酵母单杂交试验表明,苹果BR信号通路的核心转录因子MdBZR1能够特异性结合MdDWF4基因启动子区域,这表明MdBZR1通过调控MdDWF4的表达影响内源BL的生物合成。这一发现揭示了在苹果中存在一个保守的BR信号反馈调控回路:即BR信号激活后,MdBZR1不仅调控下游应答基因,还通过正向调控MdDWF4的表达来放大BR合成信号,从而精细调控内源BR水平以应对发育和环境需求。这为解释盐胁迫下MdDWF4表达上调提供了潜在的分子机制。然而,MdDWF4调控抗逆性的具体分子机制,特别是其与脱落酸(Abscisic acid,ABA)合成、ROS稳态以及植物形态建成-胁迫耐受性互作的关系有待深入研究。
综上,本研究为利用BR信号通路协同苹果抗逆性提供了理论依据。未来工作将聚焦于解析MdDWF4的具体分子调控网络,以推动耐盐苹果品种的定向育种。
-
本研究鉴定并克隆了柱型苹果中参与BR合成的关键基因MdDWF4,其蛋白结构保守且定位于内质网;功能试验表明MdDWF4过表达可提高转基因拟南芥的盐胁迫抗逆性。本研究证实了该基因通过BR通路协同参与苹果的非生物胁迫响应,为苹果耐逆育种提供了关键基因与理论依据。