



 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
光叶菝葜(Smilax glabra Roxb.)为菝葜科(Smilacaceae)菝葜属(Smilax)多年生藤本植物,其干燥根茎入药称“土茯苓”,在中医药中应用广泛。据报道该植物在全球分布于17个国家,在我国主要集中于南方省份,如广东、广西、湖南、湖北、四川及浙江等地[1],除药用外,也常用于食品和茶饮的生产[2]。光叶菝葜喜温暖湿润和荫蔽环境,多生长于海拔2 000 m以下的山坡或林下。在民族地区,光叶菝葜长期作为民间药物使用,常用于妇科炎症、风湿痹痛及梅毒等疾病的防治,深得民众信赖[3]。现代药理研究显示,光叶菝葜富含黄酮类、酚酸类和皂苷类等多种生物活性成分[2]。研究还发现,其中茎、叶的黄酮含量显著高于根茎,叶中黄酮含量可达根茎的12倍[4],表明其地上部分同样具备良好的药用与开发价值。
在广西,光叶菝葜作为一种重要药用资源,于南宁、钦州等地广泛种植。然而,在种植过程中常受多种病害威胁,主要包括炭疽病、叶斑病和锈病等[5-7],严重影响药材的产量与质量,并对当地药用植物产业的可持续发展构成威胁。南宁某光叶菝葜种植基地发生的一种叶部病害,典型症状表现为叶片出现棕色圆形或不规则病斑,严重时引发叶组织坏死与早期落叶,对药材生产构成直接威胁。为此,本研究综合采用组织分离、形态学鉴定及基于多基因(ITS、SSU、LSU、TEF、GAPDH和RPB2)的系统发育分析等方法,以明确其病原菌的分类地位。在此基础上,进一步开展了植物源杀菌剂的室内筛选,以期筛选出高效、低毒的绿色防控药剂。研究成果可为光叶菝葜叶斑病的准确诊断与绿色防控提供科学依据,对保障药材安全生产具有重要意义。
全文HTML
-
主要培养基:马铃薯葡萄糖琼脂培养基(PDA)由马铃薯200 g、葡萄糖20 g、琼脂粉20 g和蒸馏水1 000 mL配制而成;燕麦琼脂培养基(OA)由燕麦片30 g、琼脂粉20 g和蒸馏水1 000 mL配制;麦芽浸膏琼脂培养基(MEA)由麦芽浸膏30 g、琼脂粉20 g和蒸馏水1 000 mL配制;查氏琼脂培养基(CZA)由KNO3 1 g、MgSO4 0.25 g、KCl 0.5 g、葡萄糖0.2 g、蔗糖0.2 g、琼脂粉15 g和蒸馏水1 000 mL配制。
主要试剂包括CTAB(十六烷基三甲基溴化铵)、异戊醇、75%乙醇、三氯甲烷、液氮等。PCR及电泳相关试剂包括普通DNA Taq聚合酶、10×EasyTaq Buffer、DNA Marker、GelRed、6×Loading Buffer、dNTP、TBE、琼脂糖、EDTA等。
-
2022年5月在广西钦州光叶菝葜种植区发现一种叶斑病后,选取具有典型症状的罹病植株,同时移栽健康植株作为对照,共同置于广西民族大学试验大棚(温度25~32 ℃,相对湿度75%~90%,自然光照)进行培养。在统一管理及定期浇水的条件下,持续观察并记录植株的发病进程。从中采集典型叶斑病样本,用于后续病原菌的分离与鉴定。
-
病原菌的分离参照常规组织分离法[8]进行。选取光叶菝葜发病植株叶片,清洗晾干后,切取病健交界处5 mm×5 mm组织块作为分离材料。在超净工作台中,依次采用75%乙醇处理30 s、1%次氯酸钠溶液表面消毒3 min,随后用无菌水漂洗3次以去除残留消毒剂。将表面消毒后的组织块接种至含25 μg/mL氯霉素的PDA平板,于28 ℃黑暗条件下倒置培养。待菌丝萌发后,挑取菌落边缘新鲜菌丝转接至新的PDA平板进行继代培养。进一步采用单孢分离法纯化菌株:用无菌水洗脱培养7 d的菌落孢子,制备孢子悬液,经血球计数板调整浓度至1个孢子/μL,取2 μL进行平板划线,在显微镜下定位单个萌发孢子并挑取至新PDA平板,获得纯培养物,并于4 ℃条件下采用PDA斜面保存备用。
-
将分离纯化的菌株接种于含25 μg/mL氯霉素的PDA平板,28 ℃培养3~5 d备用。选取健康光叶菝葜植株,用75%乙醇擦拭消毒叶片,无菌水冲洗3次后用于致病性测定。采用刺伤接种法:用5 mm打孔器制取菌丝块,在叶片上以无菌针刺伤后覆盖菌块,以空白PDA培养基块作为对照。接种后用保鲜袋包裹保湿,定期观察并记录发病情况。发病后重新分离病原菌并按柯赫氏法则验证。
-
将供试菌株分别接种于PDA、OA、MEA平板中央,置于28 ℃黑暗条件下培养。培养过程中测量菌落直径,待菌落长满培养基时,记录菌落颜色、质地及培养基背面色泽变化,并拍照保存。无菌条件下,用接种针挑取少量菌丝于载玻片中央,加1滴无菌蒸馏水制成临时装片,在光学显微镜下观察分生孢子形态、大小及产孢细胞等特征,对典型结构显微拍照,并用ImageJ 1.53软件测量其尺寸。参照Woudenberg等[9]的方法进行形态学初步鉴定。
-
将纯培养菌株接种于PDA培养基,28 ℃暗培养7 d,用无菌牙签刮取新鲜菌丝,用CTAB法[10]提取基因组DNA。以该DNA为模板,采用表 1所列引物对ITS、SSU、LSU、RPB2、TEF和GAPDH片段进行PCR扩增(所有引物均由武汉奥科鼎盛生物科技有限公司合成)。PCR体系(25 μL):DNA模板1 μL(约50 ng)、10× EasyTaq Buffer 2.5 μL、dNTP混合液(各2.5 mM)0.5 μL、上游及下游引物(10 μM)各0.5 μL、EasyTaq DNA聚合酶(5 U/μL)0.1 μL,无菌去离子水补至25 μL。PCR反应程序:94 ℃预变性3 min;35个循环(94 ℃ 45 s,55 ℃ 30 s,72 ℃ 45 s);72 ℃终延伸10 min,4 ℃保存。PCR产物经1.2%琼脂糖凝胶电泳检测后,送公司双向测序。使用Vector NTI软件对测序峰图进行校对与拼接。将所得序列在NCBI数据库中进行BLAST同源性比对,并将最终确认的序列提交至GenBank获取登录号(表 2)。
参照Woudenberg等[9]的方法,从NCBI下载47株近缘菌株及外类群Cicatricea salina CBS 302.84的ITS、SSU、LSU、RPB2、TEF和GAPDH序列。使用PhyloSuite v1.2.2软件[17]进行如下处理:首先将各基因位点的序列分别采用MAFFT模块进行比对,然后通过Gblocks模块去除模糊比对区域(保留保守位点),最后使用concatenated sequence模块将不同基因的比对序列拼接成超级矩阵。利用PartitionFinder模块筛选最优的分区方案和核苷酸替代模型。基于处理后的序列数据集,采用IQ-TREE软件构建最大似然(Maximum Likelihood,ML)系统发育树,自举检验(Bootstrap)重复次数为20 000次。同时,使用MrBayes模块进行贝叶斯推断(Bayesian Inference,BI),运行4条马尔可夫链,每1 000代采样一次,直至分裂频率标准差低于0.01。最终,使用FigTree软件对系统发育树进行可视化与注释。
-
选用和厚朴酚(麦克林,纯度≥98%,货号M813634)、柠檬醛(麦克林,纯度97%,货号C804526)、香茅醛(麦克林,纯度96%,货号C805140)、香叶醇(麦克林,纯度98%,货号G810439)、麝香草酚(麦克林,纯度98%,货号T818893)和左旋香芹酮(麦克林,纯度99%,货号L805683)6种植物源杀菌剂进行室内毒力测定。根据各化合物的溶解特性,先配制成50~100 mg/mL母液,再用无菌PDA培养基逐级稀释至预试确定的浓度梯度,并添加终浓度25 μg/mL氯霉素以抑制细菌污染。将药液与预冷至约50 ℃的PDA充分混匀后倒板,凝固制成含药平板。确定杀菌剂浓度[18]及前期预试验,设置5个浓度梯度。用5 mm无菌打孔器切取菌落边缘菌丝块,菌丝面朝下接种于含药平板中央,每浓度3次重复,以无药PDA为空白对照。接种皿用封口膜密封,28 ℃倒置培养5 d后,采用十字交叉法测量各处理的菌落直径,并按以下公式计算菌丝生长抑制率。
式中:0.5 cm为接种菌丝块的直径。
-
试验数据整理后,计算平均值与标准差。采用GraphPad Prism 8.0软件进行统计分析。采用单因素方差分析(One-Way ANOVA)比较组间差异,若存在显著性(p<0.05),则进一步使用Tukey法进行多重比较。采用非线性回归模型进行毒力回归分析,计算各药剂的半数有效浓度(EC50)、95%置信区间(95% CI)、斜率及决定系数(R2)。试验结果均以平均值±标准差表示,并采用柱状图与折线图进行可视化。
1.1. 供试培养基及试剂
1.2. 病样采集
1.3. 病原菌分离、纯化及保存
1.4. 致病性试验
1.5. 形态学观察
1.6. 分子生物学鉴定
1.7. 不同杀菌剂的室内毒力测定
1.8. 统计分析
-
2022年5月,在广西南宁光叶菝葜种植基地发现叶斑病为害。病害初期表现为直径1~2 mm的深褐色圆形小斑,边缘清晰;随着病情发展,病斑数量增多并逐渐扩展为3~5 mm的圆形或近圆形斑块,颜色由浅棕色转为深棕色(图 1)。在高湿(相对湿度>90%)环境下,多个病斑相互融合,造成叶片大面积枯死,可见深棕色轮纹,严重时叶片出现萎蔫或脱落现象。
-
通过组织分离法共获得9株真菌分离物,其菌落形态特征基本一致。经叶片接种测定,所有菌株均能在光叶菝葜上引起叶斑症状。选取代表性菌株TFL18.1进行单孢纯化后用于后续致病性研究。采用菌丝块接种法进行回接试验,将直径5 mm的菌丝块贴接于经表面刺伤的健株叶片上。接种第3 d,叶片开始出现规则的圆形浅色褪绿病斑(图 2b、2c);至第6 d,病斑持续扩展,形成棕黄色坏死斑,表面产生灰白色霉状物(图 2e、2f),其症状特征与田间观察结果类似。空白对照叶片在整个试验期间无病斑(图 2a)。从发病组织重新分离获得的菌株,其形态特征与原始菌株TFL18.1一致,符合柯赫氏法则的验证要求,据此确认菌株TFL18.1为光叶菝葜叶斑病的致病菌。
-
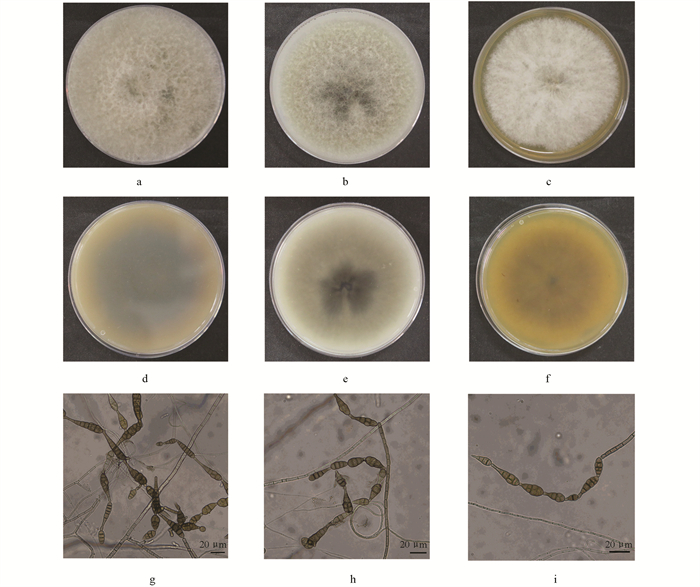
将菌株TFL18.1分别接种于PDA、OA和MEA培养基,于28 ℃黑暗条件下培养6 d后,观察其菌落及显微形态特征。在PDA培养基上,菌落扩展迅速,直径达9 cm,覆盖整个培养皿;气生菌丝呈白色且生长丰密,菌落正面无明显色素产生(图 3a),背面中心为黑色,边缘呈浅棕色(图 3d)。在OA培养基上,菌落呈规则圆形;气生菌丝茂密、白色,中心区域呈灰黑色(图 3b);背面中心为深灰色,向外渐变为淡灰褐色,边缘为白色(图 3e)。在MEA培养基上,菌落为圆形,气生菌丝致密、呈绒毛状,白色(图 3c);背面中心为土黄色,向边缘逐渐过渡为浅黄色(图 3f)。显微结构观察表明,分生孢子梗单生或簇生,直立或微弯曲,分枝较少;分生孢子呈棕色至褐色,形态多样,包括棒状、梨形或椭圆锥形,具横隔膜与纵隔膜,隔膜处略缢缩,孢子大小为(19~34) μm × (3.5~12.5)μm(图 3g、3h、3i)。综合上述菌落形态与孢子显微特征,菌株TFL18.1符合链格孢属(Alternaria)[9]的典型特征,初步鉴定为一种链格孢菌(Alternaria sp.)。
-
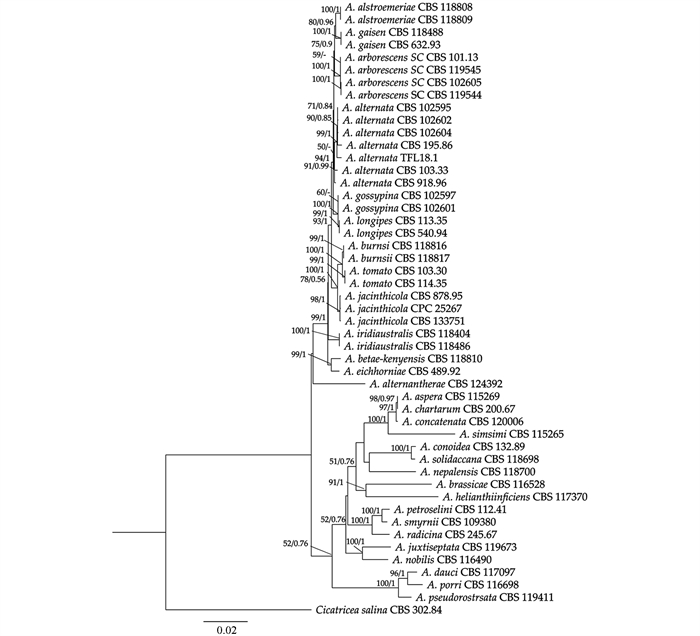
为准确鉴定菌株TFL18.1的分类地位,本研究选用引物对ITS-1/ITS-4、NS1/NS4、LROR/LR5、EF-983F/EF1-2128R、GPD-1/GPD-2及RPB2-5F/fRPB2-7cR,以其基因组DNA为模板,分别扩增ITS、SSU、LSU、TEF、GAPDH和RPB2基因片段,各PCR产物经纯化后送至测序公司进行双向测序,经拼接和校对后的长度分别为:579、1 056、959、242、626和750 bp,提交至NCBI GenBank分别获得登录号ITS (PP292012)、SSU (PP292027)、LSU (PP292026)、TEF (PP318628)、GAPDH (PP318627)、RPB2 (PP318626)。BLASTn单基因比对分析显示,6个片段与Alternaria alternata CBS 195.86的相似度分别为100%(ITS,515/515 bp)、99%(SSU,1 020/1 021 bp)、99%(LSU,849/850 bp)、100%(TEF,240/240 bp)、99%(GAPDH,572/579 bp)和99%(RPB2,732/736 bp)。进一步将上述6个基因序列进行串联,分别采用最大似然法(ML)和贝叶斯推断法(BI)构建系统发育树(图 4)。结果显示,菌株TFL18.1与A. alternata典型菌株聚于同一分支,且该分支获得较高的统计支持(ML Bootstrap值=99%,BI后验概率=1.00)。综合形态学特征与多基因系统发育分析结果,确定引起光叶菝葜叶斑病的病原菌为A. alternata。
-
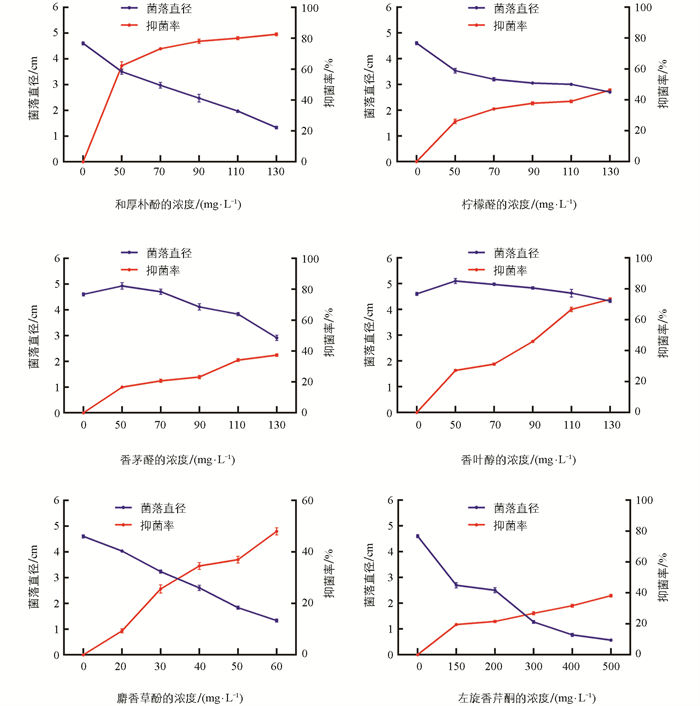
采用菌丝生长速率法测定6种植物源药剂对病原菌TFL18.1的抑制效果。结果表明,各药剂对菌丝生长的抑制效果存在显著差异,且抑制率均随浓度升高而增加(图 5)。在供试药剂中,和厚朴酚表现出最强的抑制活性,在50 mg/L浓度下抑制率达62.23%,130 mg/L时增至82.52%。麝香草酚的抑菌效果次之,60 mg/L浓度时抑制率为47.96%。香叶醇在较高浓度下也显示出良好效果,900 mg/L时抑制率为73.17%。相比之下,柠檬醛与左旋香芹酮的抑菌能力较弱,在最高测试浓度(分别为600 mg/L和500 mg/L)下,抑制率仅为46.33%和38.21%;香茅醛的效果最差,900 mg/L时抑制率仅37.40%。
毒力回归分析显示,6种药剂的毒力回归方程拟合良好(R2=0.878 7~0.945 5)。和厚朴酚的毒力最强,EC50值为45.56 mg/L;麝香草酚次之,EC50值为63.69 mg/L。香茅醛毒力中等,EC50值为462.82 mg/L。香叶醇、左旋香芹酮和柠檬醛的毒力相对较弱,EC50值分别为687.30、994.72和2 084.00 mg/L。各药剂毒力从大到小依次为:和厚朴酚、麝香草酚、香茅醛、香叶醇、左旋香芹酮、柠檬醛(表 3)。综上,和厚朴酚和麝香草酚对光叶菝葜叶斑病病原菌表现出显著的抑制效果,具备作为该病害潜在绿色防控药剂的开发价值。
2.1. 病害症状
2.2. 病原分离与致病性测定
2.3. 病原菌形态学鉴定
2.4. 分子生物学鉴定
2.5. 不同植物源药剂的室内毒力测定
-
链格孢属(Alternaria)隶属于真菌界子囊菌门(Ascomycota)、座囊菌纲(Dothideomycetes)、格孢腔菌目(Pleosporales)、格孢腔菌科(Pleosporaceae)[19],广泛分布于全球多种生态环境中,包括寄生、腐生和内生类群,可侵染多种作物及药用植物,引致叶斑、果腐和茎腐等病害,造成严重经济损失[20-21]。Woudenberg等[9]依据多基因系统发育关系将该属划分为28个组,其中A. alternata作为模式种被归入Alternaria组,该组目前包含11个种和1个复合种[22]。本研究通过致病性验证、形态特征及多基因(ITS、SSU、LSU、TEF、GAPDH、RPB2)联合系统发育分析,明确了A. alternata为光叶菝葜(Smilax glabra)叶斑病的病原菌,扩展了该病原的寄主范围,为病害诊断与防控提供了依据。然而,本研究样本仅来源于广西南宁和钦州的种植基地,其结果可能难以代表该病害在不同生态区的流行规律和病原遗传多样性。今后需开展更大范围的病原收集(如覆盖华南地区主要种植区)与群体遗传结构分析,并结合基因组数据深入解析其致病机制。关于A. alternata的致病机制,现有研究表明其能产生多种寄主选择性毒素(Host-Specific toxins,HSTs),如ACT-毒素、AF-毒素等,这些毒素可导致植物细胞质膜透性改变、电解质渗漏,最终引起细胞坏死[20]。本研究中观察到的叶片坏死症状可能与该菌产生相关毒素有关,后续需进一步从代谢产物层面解析其侵染机理。
植物病原真菌是为害植物的重要病原,超过70%的植物病害由真菌引起,对作物产量和品质构成严重威胁。虽然化学农药是目前最有效的防治手段,但长期大量使用已带来环境安全、人畜健康等诸多问题[23]。相较于针对链格孢属病害的高效化学杀菌剂(如咯菌腈、嘧菌环胺、咪鲜胺等,其EC50值通常<1 mg/L)[24-26],本研究筛选出的和厚朴酚与麝香草酚的EC50值(分别为45.56 mg/L和63.69 mg/L)相对较高。然而,鉴于链格孢菌已对多种化学药剂(如啶酰菌胺、甲氧基丙烯酸酯类)产生严重的抗药性[24-25],且植物源杀菌剂具有易降解、环境残留低及作用机制多样等优势,它们在病害综合治理及抗药性管理中仍具有重要的开发潜力。近年来,植物源活性成分因其广谱抗菌活性、环境友好性和较低毒性[27],在植物病害绿色防控领域备受关注,已成为新农药研发的重要方向[28]。诸如黄酮类、萜类和酚类等植物次生代谢产物,在微生物防控中通常具备多靶点作用机制,不易诱导抗药性[25]。和厚朴酚与麝香草酚均为酚类或酚类衍生物,其抑菌机制可能涉及破坏病原菌细胞膜的完整性,导致细胞内含物外泄,或干扰线粒体呼吸链,抑制能量代谢[29]。研究表明,石荠苎和红梗楠的丙酮提取物对茄链格孢菌(Alternaria solani)、茄腐镰刀菌(Fusarium solani)、香蕉炭疽刺盘孢菌(Colletotrichum musae)和水稻纹枯病菌(Rhizoctonia solani)4种植物病原真菌均有抑制作用;其中,石荠苎提取物的抑菌率为70.2%~100%,红梗楠提取物的抑菌率为50.6%~92.8%[26]。另有研究采用菌落生长速率法测定7种植物化学物质对果生刺盘孢(Colletotrichum fructicola)的抑制效果,发现和厚朴酚、厚朴酚、麝香草酚和香芹酚均表现出良好的抗真菌活性,其中和厚朴酚与厚朴酚在田间对果生刺盘孢引起的炭疽病也表现出良好的防治效果[27]。本研究中,和厚朴酚与麝香草酚在室内条件下对光叶菝葜叶斑病原菌A. alternata表现出显著的抑制效果。然而,本研究目前仅局限于室内毒力筛选,尚未开展田间应用效果验证。后续需通过田间小区试验及大田示范,进一步评估该类药剂的实际防效与环境相容性;并应深入解析其抑菌作用机理,为药用植物病害的绿色可持续治理提供更为充分的科学依据。
-
本研究首次证实引起光叶菝葜(Smilax glabra)叶斑病的病原菌为链格孢(Alternaria alternata),同时筛选出两种对致病菌株TFL18.1具有显著抑菌作用的高效植物源杀菌剂:和厚朴酚和麝香草酚。本研究结果不仅明确了病原菌分类地位,也为该病害的绿色防控提供了新选择,对促进光叶菝葜产业的健康可持续发展具有重要指导意义。