


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
土壤微生物的组成、多样性和功能直接参与土壤养分循环、有机质分解、植物生长调控及生态系统稳定性维持等关键过程[1]。农林复合指在同一块土地上先种植一种作物,然后再种植另一种作物的种植方式,这种方式有助于充分利用土地资源并提高作物生产力[2],对于增加土壤微生物的多样性[3]和促进植物生长起重要作用[4]。土壤微生物一般以根系分泌物和植物根、茎、枝、叶为营养物质,植物种类不同对土壤碳氮循环的影响存在差异[5],种植不同植物可改变土壤微生物群落的结构和功能[6]。不同的植物组成会引发不同的种间作用,起到相互促进或抑制的效应,这种相互作用通过土壤和微生物的调节活动间接发生[7]。适宜的农林复合模式不仅能够促进植物根系对土壤养分的吸收和利用,促进植物快速生长[8],还可促使植物根际土壤微生物的生物量和群落结构丰富度增加,使土壤微生物多样性显著增加[9]。这对农林生产有较大帮助,如加速土壤间养分循环、提高林木抗性、修复生态环境以及调节土壤结构等。
薄壳山核桃(Carya illinoinensis)作为一种兼具经济与生态价值的高效经济树种,近年来在我国南方地区被广泛推广种植。为提高土地资源利用效率、改善果园生态环境,套种模式逐渐受到关注。豆科牧草通过其根瘤固氮作用能够促进固氮菌和共生菌的生长,也能够通过其化感作用影响其他植物的生长[10],牧草间作可通过促进根系分泌糖类、酚酸类等多种物质为微生物群落提供丰富的碳源和能量,从而改变微生物的多样性和群落结构[11]。季琳琳等[12]对山核桃间作油菜、绵枣儿、野豌豆进行了研究,表明不同草类对林地土壤微生物群落结构影响的差异有统计学意义:各生草处理0~10 cm土壤中球囊菌门相对丰度较对照组分别提高202.81%、193.90%、65.04%;间作油菜能有效改善山核桃林地微生物群落结构,提高土壤细菌群落的物种丰富度和均匀度。这对于修复退化山核桃林地和优化林地土壤环境具有重要参考意义。何升然等[13]研究表明,相比单作,间作紫花苜蓿能有效提高甜高粱根际土壤中拟杆菌门、放线菌门、变形菌门和Gp6的相对丰度,其中变形菌门和拟杆菌门对根际土壤中碳、氮等养分起积极作用,表明在甜高粱地间作紫花苜蓿有利于土壤微生态环境的改善。鲍佳书等[14]分析比较了不同品种薄壳山核桃土壤微生物的多样性及群落组成。刘丽丽等[15]研究了薄壳山核桃根际与非根际土壤微生物群落结构特征。张嘉恬等[16]研究了薄壳山核桃套种小麦土壤细菌及真菌的多样性、群落结构、标志物种和共现网络关系。关于薄壳山核桃套种苜蓿和将军菊苣对土壤微生物群落的调控效应尚未有相关报道。本研究以薄壳山核桃与苜蓿、将军菊苣的套种系统作为研究对象,采用高通量测序技术,探讨不同牧草套种模式下土壤微生物群落结构和多样性变化,以及微生物群落与土壤环境因子间的关系,旨在为优化薄壳山核桃复合种植模式、提升土壤生态服务功能提供理论依据,这对于深入了解薄壳山核桃套种不同牧草与土壤微生物之间的复杂关系、实现果园的可持续健康经营具有重要意义。
全文HTML
-
实验地位于湖北省京山市虎爪山林场,处于鄂中丘陵至江汉平原的过渡地带(30°42′-31°27′N,112°43′-113°29′E)。属北亚热带季风气候区,年平均气温16.3 ℃,极端最高气温40.3 ℃,极端最低气温-6.2 ℃。无霜期223~243 d,年均太阳辐射总量4 520~4 600 MJ/m2,日照总时数1 970.5 h。年平均降水量1 170 mm,夏季降水量约占全年的41%~45%,冬季占7%~8%,春季占28%~32%,秋季占18%~20%,年降水天数104 ~130 d。土壤主要为沙壤,土层深厚,透气性良好,水解氮含量102 mg/kg,有效磷含量20 mg/kg,速效钾含量186 mg/kg,有机质含量17.1 g/kg,pH值5.52。
-
2022年3月,在造林5年的薄壳山核桃林地套种苜蓿和将军菊苣,薄壳山核桃林株行距为8 m×8 m,平均树高3.5 m,冠幅3.1 m×2.8 m。实验设计3种种植模式,分别为薄壳山核桃+苜蓿、薄壳山核桃+将军菊苣,以及薄壳山核桃单作不套种。苜蓿和将军菊苣均采用种子撒播,播种时,除以薄壳山核桃树干为中心、长宽均为2 m矩形范围内不撒种外,其余地方全播种,播种量均为37.5 kg/hm2,3次重复。每年6月和11月对将军菊苣共刈割2次,苜蓿不刈割。
-
土壤样品于2024年8月28日采集,每种模式每次重复均选取3株薄壳山核桃,分别在根际和非根际范围内采集0~20 cm深度土壤混合,清除石砾、根系及杂物。采用四分法,取1/4土壤作为1份样品。套种苜蓿和将军菊苣模式的根际和非根际土样分别采集样品3份,单作模式采集根际土样3份。根际土壤为距树干50 cm范围内土样,其他为非根际土壤。本研究划分5类土样:套种苜蓿的薄壳山核桃根际土样记为MMG,套种苜蓿的薄壳山核桃非根际土样记为MMCG,套种将军菊苣的薄壳山核桃根际土样记为MJG,套种将军菊苣的薄壳山核桃非根际土样记为MJCG,薄壳山核桃单作根际土样记为MCG,共15份。将每份土壤样品分成2份,过筛去除杂物后放入冰盒带回实验室,1份用于土壤微生物高通量测序,另1份自然风干,粉碎过60目筛后用于土壤理化性状测定。
-
将0.5 g新鲜土壤加入到盛有提取裂解液的离心管中进行研磨处理,采用OMEGA Soil DNA Kit试剂盒提取核酸,以0.8%琼脂糖凝胶电泳检测提取土壤的微生物DNA总质量,利用Nanodrop 2000对DNA进行定量。细菌使用引物341F/806R对16S基因的V3-V4区域进行扩增,引物序列为341F(CCTAYGGGRBGCASCAG)和806R(GGACTACNNGGGTATCTAAT)。真菌采用通用引物ITS1F(CTTGGTCATTTAGAGGAAGTAA)和ITS2R(GCTGCGTTCTTCATCGATGC)对ITS基因的ITS1区域进行扩增。16S和ITS扩增分别采用TransStart Fastpfu DNA聚合酶、TaKaRa rTaq DNA聚合酶和20 μL反应体系。PCR反应条件及扩增程序为:94 ℃预变性2 min,细菌30个循环(94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),真菌30个循环(94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),最后在72 ℃延伸2 min,于4 ℃保存。扩增结束后,回收PCR产物,经过定量后,利用Illumina Miseq PE300平台进行测序,原始测序序列下机后,测序公司进行初步质控,并发回高质量测序序列,由武汉爱基百客生物科技有限公司完成。
-
首先使用Flash软件对序列进行拼接,使用UPARSE对序列进行OTU聚类,相似度设为97%,聚类过程中去除嵌合体,获得OTU代表序列。群落结构分析等内容需要使用重抽后的OTU表,重抽序列阈值为75 825。使用RDP classifier对代表性聚类进行分类学注释,使用数据库为Silva 16S rRNA数据库(v138),比对阈值为75%。计算土壤微生物α多样性,分析细菌与真菌群落的结构组成及其与土壤水解氮(Hydrolyzable nitrogen,AN)、有效磷(Available phosphorus,AP)、速效钾(Instant Potassium,AK)、有机质(Organic matter,AM)、pH值、脲酶(Urease,URE)、酸性磷酸酶(Acid phosphatase,APH)及蔗糖酶(Sucrase,SUC)等环境因子的相关性。
-
土壤pH值测定采用pH计,有机质含量测定采用重铬酸钾-硫酸稀释预热法,水解氮含量测定采用碱解扩散法,有效磷含量测定采用钼锑抗比色法,速效钾含量测定采用火焰光度法,脲酶活性测定采用硝基酚比色法,蔗糖酶活性测定采用3,5-二硝基水杨酸比色法,酸性磷酸酶活性测定采用磷酸苯二钠比色法[17]。
2.1. 实验材料与设计
2.2. 土壤样品采集
2.3. 土壤细菌16S和真菌ITS高通量测序及数据分析
2.4. 数据生信分析
2.5. 土壤理化性质测定
-
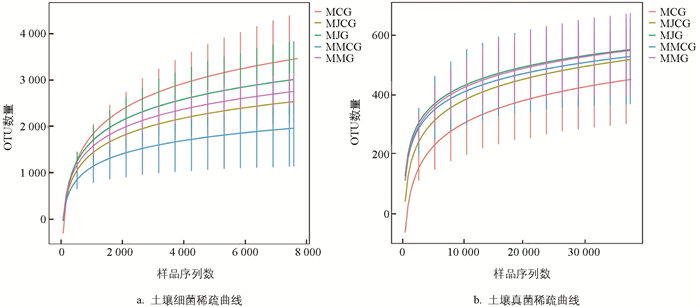
由图 1可知,当细菌OTU数量达到3 000、真菌OTU数量达到400后,稀疏曲线均趋于平稳,说明此时测序数量已达到饱和,能够反映土壤样品的微生物群落组成,测序数据合理。
-
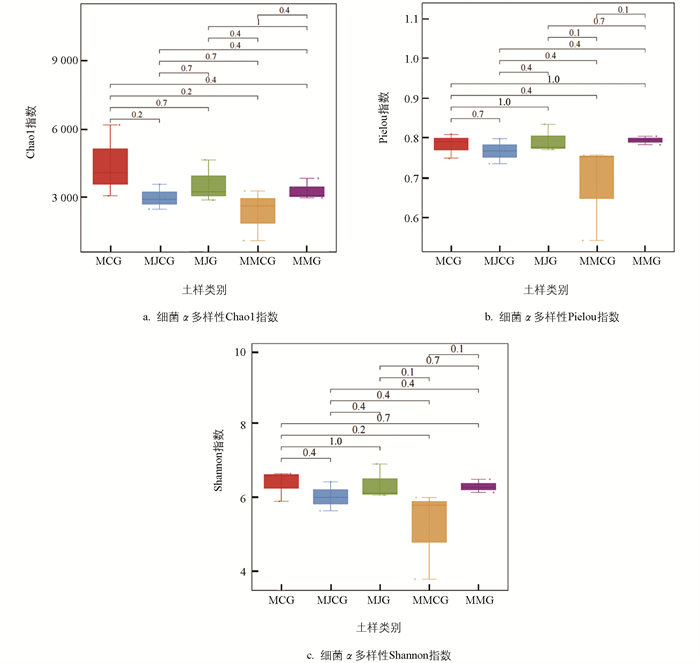
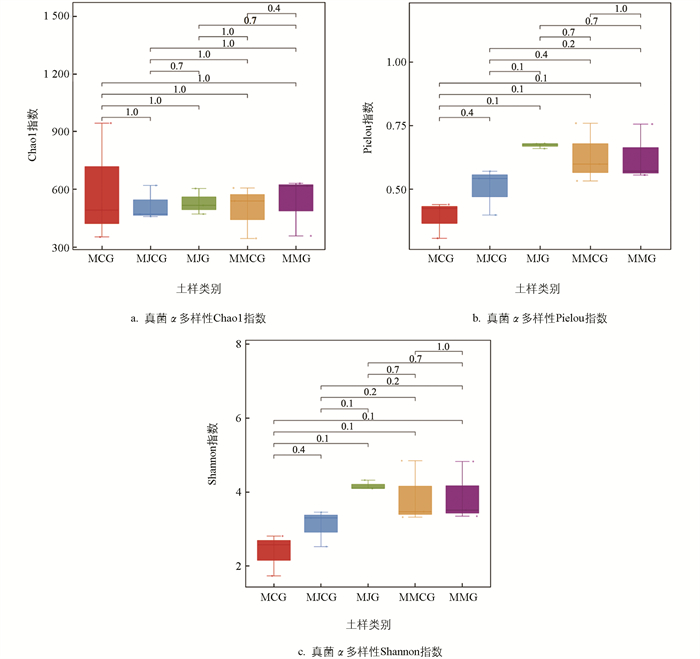
对薄壳山核桃套种苜蓿和将军菊苣模式,以及单作模式的根际、非根际土壤细菌和真菌α多样性进行分析。由图 2、图 3可知,3种模式土壤细菌和真菌α多样性Chao1指数、Pielou指数和Shannon指数的差异均无统计学意义。土壤细菌α多样性Chao1指数、Pielou指数、Shannon指数均以MCG相对较高,MJG和MMG均相对高于MJCG和MMCG;土壤真菌α多样性Chao1指数、Pielou指数、Shannon指数MJG和MMG相对高于MJCG和MMCG。表明根际土壤细菌和真菌的α多样性指数均高于非根际土壤。
-
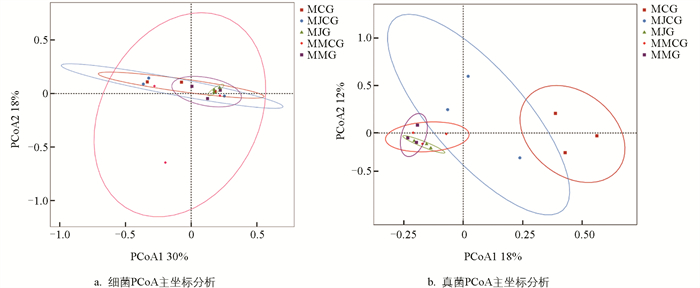
PCoA主坐标分析是一种展示样本间相似性的分析方法,与PCA分析思路基本一致,都是通过降维方式寻找复杂样本中主要样本的差异距离。与PCA不同的是,PCoA主要利用Unifrac、Bray-Curtis等样本距离信息进行计算以及降维图形展示,因此结果更集中于体现样本间的相异性距离。针对细菌群落进行PCoA主坐标分析(图 4a),PCoA1、PCoA2分别解释了30%和18%的变异,累积贡献率达48%,其中PCoA1、PCoA2是PCoA降维后的两个核心坐标轴,分别捕捉数据的主要变异方向。5类土样重叠度较高,不同样本3个重复并不位于同一象限,表明组内细菌群落差异较大,尤其以MMCG组内差异较大,组间没有发生明显的变化。针对真菌群落进行PCoA主坐标分析(图 4b),PCoA1、PCoA2分别解释了18%和12%的变异,累积贡献率达30%,MMCG和MJG位于第三象限,与其他土样真菌群落差异较大;MCG与MMG、MMCG、MJG分别位于PCoA1正负两端,真菌群落结构有明显差异;MMJG组内差异较大,MJG、MMG和MMCG组内差异较小。
-
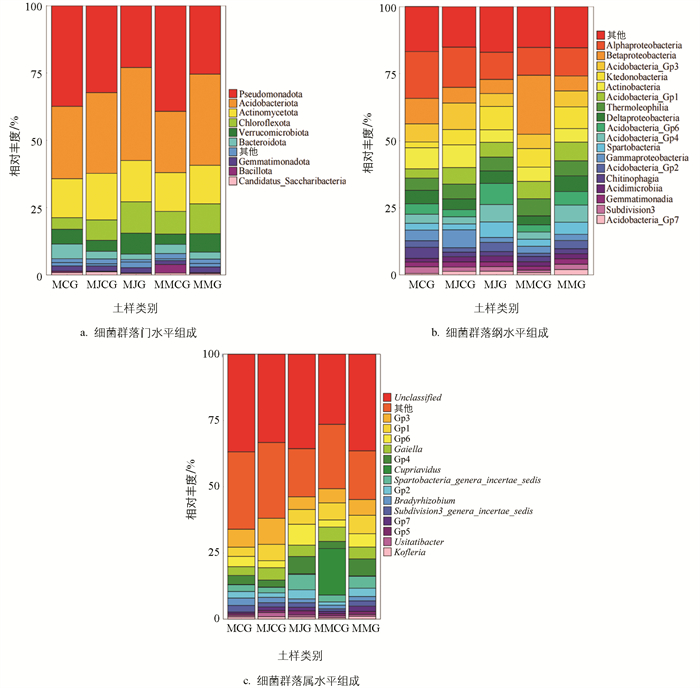
根据OUTs代表序列的注释结果,5类土样细菌群落由32门、82纲、614属组成。由图 5a可知,门水平优势菌门(相对丰度大于1%)为假单胞菌门(Pseudomonadota)、酸杆菌门(Acidobacteriota)、放线菌门(Actinomycetota)、绿弯菌门(Chloroflexota)、疣微菌门(Verrucomicrobiota)、拟杆菌门(Bacteroidota)、芽单胞菌门(Gemmatimonadota)、芽孢杆菌门(Bacillota)、Candidatus_Saccharibacteria。不同土样各菌门相对丰度存在一定差异:均以Pseudomonadota、Acidobacteriota、Actinomycetota、Chloroflexota和Bacteroidota菌门的相对丰度较大;Pseudomonadota和Bacteroidota在MJG和MMG(根际土样)中相对丰度均低于非根际土样和单作土样;Acidobacteriota、Verrucomicrobiota和Chloroflexota在MJG和MMG(根际土样)中相对丰度均高于非根际土样和单作土样;Actinomycetota在MJG和MMG(根际土样)中相对丰度均低于非根际土样。此外,在5类土样中均存在相对丰度小于1%的亚硝化球菌门(Nitrososphaerota)、硝化螺旋菌门(Nitrospirota)。
由图 5b可知,细菌纲水平优势菌纲(相对丰度大于1%)为α-变形菌纲(Alphaproteobacteria)、酸杆菌纲(Acidobacteria_Gp6,Gp6)、β-变形菌纲(Betaproteobacteria)、放线菌纲(Actinobacteria)、酸杆菌纲(Acidobacteria_Gp4,Gp4)、热嗜菌纲(Thermoleophilia)、δ-变形菌纲(Deltaproteobacteria)、酸杆菌纲Gp3(Acidobacteria_Gp3)、γ-变形菌纲(Gammaproteobacteria)、纤线杆菌纲(Ktedonobacteria)、噬几丁质菌纲(Chitinophagia)、酸杆菌纲Gp1(Acidobacteria_Gp1)、Gemmatimonadia、斯巴杆菌纲(Spartobacteria)、酸微菌纲(Acidimicrobiia)、Subdivision3、酸杆菌纲Gp7(Acidobacteria_Gp7)、酸杆菌纲Gp2(Acidobacteria_Gp2)。不同土样各优势菌纲存在一定差异:Alphaproteobacteria、Betaproteobacteria、Acidobacteria_Gp3在MJG和MMG(根际土样)中相对丰度均低于非根际土样和单作土样;Ktedonobacteria在MJG和MMG(根际土样)中相对丰度均高于非根际土样和单作土样;Acidobacteria_Gp1、Thermoleophilia在MJG和MMG(根际土样)中相对丰度均低于非根际土样,高于单作土样。表明根际、非根际、单作间土壤细菌纲水平存在差异。
由图 5c可知,细菌属水平优势菌属(相对丰度大于1%)为Unclassified、Gp6、Gp4、Gp3、放线菌属(Gaiella)、Gp1、Spartobacteria_genera_incertae_sedis、Subdivision3_genera_incertae_sedis、贪铜菌属(Cupriavidus)、Gp7、Bradyrhizobium、Gp2、Usitatibacter、Kofleria、Gp5。不同土样各优势菌属存在一定差异:Unclassified、Gp6、Gp4、Gp2、Spartobacteria_genera_incertae_sedis在MJG和MMG(根际土样)中相对丰度均高于非根际土样和单作土样;Gp3、Gp1在MJG土样中相对丰度低于MJCG土样,在MMG土样中高于MMCG土样;Gaiella在MJG和MMG(根际土样)中相对丰度均低于非根际土样,高于单作土样;Cupriavidus在MMCG土样中相对丰度显著高于其他土样。表明根际、非根际、单作间土壤细菌属水平存在差异。此外,在5类土样中均存在亚硝化球菌属Nitrososphaera和消化螺菌属Nitrospira,相对丰度均小于1%。
-
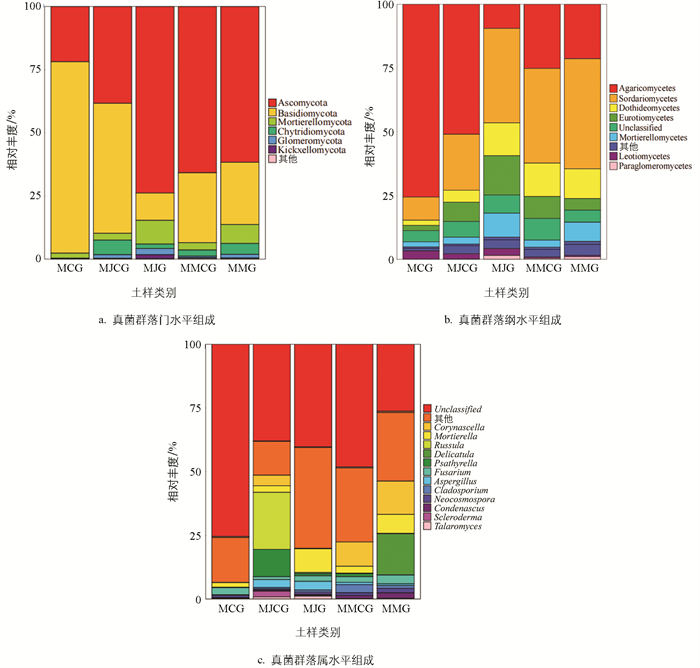
根据OTUs代表序列的注释结果,5类土样真菌群落由13门、55纲、636属组成。由图 6a可知,真菌优势菌门(相对丰度大于1%)主要有子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、被孢霉门(Mortierellomycota)、壶菌门(Chytridiomycota)、球囊菌门(Glomeromycota)、梳霉门(Kickxellomycota)。不同土样优势菌门相对丰度存在差异:Ascomycota在MJG土样中相对丰度最大,在MMG土样中相对丰度低于MMCG土样,但都显著高于单作土样;Basidiomycota在MJG土样和MMG土样中相对丰度均低于非根际土样和单作土样,单作土样最高;Mortierellomycota在MJG和MMG土样中相对丰度均显著高于非根际土样和单作土样;Chytridiomycota在套种土样中相对丰度显著高于单作土样;Glomeromycota存在于套种土样中,但在单作土样中不存在。表明真菌群落门水平存在差异,套种后发生了显著变化。
由图 6b可知,真菌优势菌纲(相对丰度大于1%)主要有粪壳菌纲(Sordariomycetes)、伞菌纲(Agaricomycetes)、座囊菌纲(Dothideomycetes)、被孢霉纲(Mortierellomycetes)、散囊菌纲(Eurotiomycetes)、Unclassified、锤舌菌纲(Leotiomycetes)、类球囊霉纲(Paraglomeromycetes)。其中:Agaricomycetes在单作土样中相对丰度显著高于套种土样,在根际土样中的相对丰度小于非根际土样;Sordariomycetes在套种土样中相对丰度高于单作土样,且根际土样中相对丰度大于非根际土样;Dothideomycetes、Eurotiomycetes、Unclassified和Mortierellomycetes在套种土样中相对丰度高于单作土样;Paraglomeromycetes在单作土样中不存在,而在套种根际土样和非根际土样中均存在。表明套种后真菌群落纲水平发生了显著变化。
由图 6c可知,真菌优势菌属(相对丰度大于1%)主要有Unclassified、被孢霉属(Mortierella)、硬皮马勃属(Scleroderma)、Corynascella、脆柄菇属(Psathyrella)、红菇属(Russula)、Condenascus、曲霉菌属(Aspergillus)、Delicatula、新球壳孢属(Neocosmospora)、枝孢属(Cladosporium)、篮状菌属(Talaromyces)。其中:Unclassified在单作土样中相对丰度大于套种土样;Corynascella在套种土样中相对丰度大于单作土样,且在根际土样中大于非根际土样;根据微生物测定报告,在单作土样中存在Tuber,而在套种土样中不存在。表明套种对土壤微生物群落结构产生了较大影响。
-
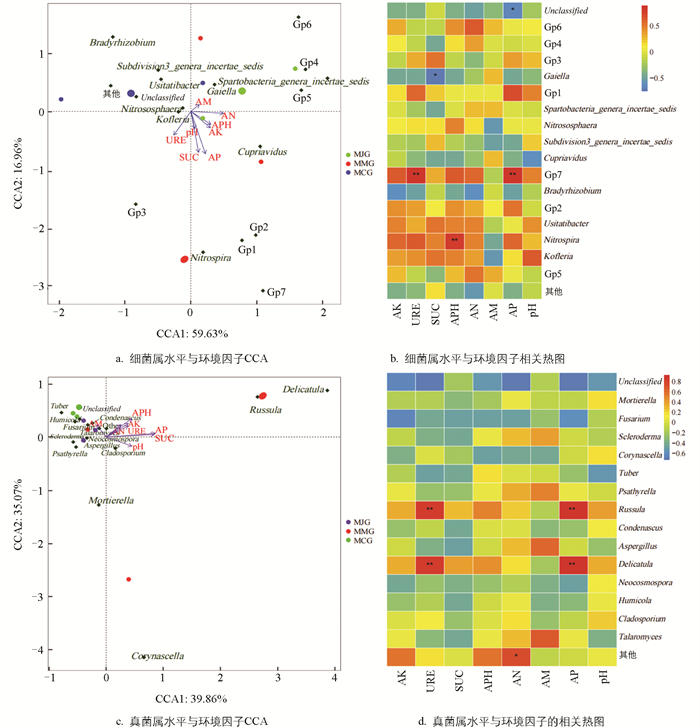
利用基于单峰模型的多元直接梯度相关分析探讨不同牧草套种模式下细菌属、真菌属与理化性状的相关关系。CCA是一种多变量统计方法,通过降维技术揭示不同环境因子与维生物群落结构之间的关联模式,CCA1、CCA2是该分析中的两种主要模式。由图 7a可知,环境因子CCA1和CCA2的累积贡献率为76.59%,表明8个环境因子(pH、AM、AN、APH、AK、AP、SUC、URE)总体对3种模式根际土壤细菌群落有显著影响,其中AP对细菌群落组成有显著影响,是细菌群落结构构建的主导因子,MJG土样细菌群落主要受AP影响。pH、SUC、AP、AK、APH和AN箭头指向第二象限,与Cupriavidus、Nitrospira、Gp2、Gp1、Gp7呈正相关。图 7b显示,AP、URE与Gp7,APH与Nitrospira呈显著正相关;SUC与Gaiella,AP与Unclassified呈显著负相关。
图 7c显示,环境因子CCA1和CCA2累积贡献率为74.93%,8个环境因子总体对3种模式根际土壤真菌群落有显著影响,其中SUC和AP对真菌群落组成有显著影响,是真菌群落结构构建的主导因子,对MJG和MCG土样真菌群落构建会产生不利影响。图 7d显示,URE、AP与Russula和Delicatula呈极显著正相关。
3.1. 土壤微生物样品测序结果
3.2. 不同套种模式的土壤微生物α多样性
3.3. 不同套种模式的土壤微生物细菌、真菌PCoA主坐标分析
3.4. 不同套种模式的土壤微生物群落结构影响
3.4.1. 不同套种模式的土壤细菌群落结构分析
3.4.2. 不同套种模式的土壤真菌群落结构分析
3.5. 土壤理化性质与微生物群落的相关性分析
-
合理的套种模式能够促进植物根际土壤细菌和真菌群落结构优化[18]。本研究中,3种模式根际与非根际土壤细菌和真菌的α多样性指数的差异均无统计学意义。不同植物根系分泌物具有一定的特异性[19],会对根际土壤微生态结构的多样性产生不同程度的影响[20],这与孙萌迪等[21]对华中五味子与玉米套种的研究结果一致。Chen等[22]在对云杉林下套种五味子的研究中也发现土壤微生物多样性差异无统计学意义的现象。Li等[23]在不同套种模式下小麦的研究中指出,土壤α多样性并未受到显著影响,与本研究结论一致。PCoA分析结果表明,MCG与MMG、MMCG、MJG土样中真菌群落结构差异较大,MJG、MMCG、MMG间真菌群落结构差异不明显,表明相比单作,套种改变了薄壳山核桃根际土壤的真菌群落结构,套种苜蓿和将军菊苣的根际土壤真菌群落结构未发生明显改变。这一现象可能的解释是物种丰度受环境影响较物种组成相对偏弱,影响通常可能发生在个别类群中,而不是整个群落[23],同时群落结构的变化并非一定会造成物种多样性的改变,因为不同真菌类群的变化会产生补偿效应[24]。
-
本研究发现,Pseudomonadota、Acidobacteriota、Actinomycetota、Chloroflexota和Bacteroidota为细菌群落优势菌门,Pseudomonadota和Bacteroidota在MJG和MMG(根际土样)中相对丰度均低于非根际和单作土样,这可能是根际区域因根系分泌物富含简单碳源,而Bacteroidota更擅长降解复杂多糖,在简单碳源主导的根际环境中竞争力较弱[25]。套种植物也可能通过释放特异性化合物抑制特定菌群,如三七生病植株根际假单胞菌富集,而健康植株通过化感作用能抑制其增殖[26];丹参套种模式下镰刀菌等病原菌减少,但Bacteroidota丰度仍受连作分泌物积累的负面影响[27]。Acidobacteriota、Verrucomicrobiota和Chloroflexota在MJG和MMG(根际土样)中相对丰度均高于非根际和单作土样;Actinomycetota在MJG和MMG(根际土样)中相对丰度均低于非根际土样。Acidobacteriota通常被认为是寡营养型细菌,适应于低营养环境,可能在有机质分解中起作用[28]。Verrucomicrobiota可能与复杂碳源(例如几丁质或纤维素)的降解有关,而Chloroflexota则包括一些光养细菌和参与有机物分解的成员,尤其是在厌氧或微氧环境中[29]。套种模式中,不同植物根系分泌物的互补性输入(如糖类、酚酸、木质素衍生物等)显著增加了根际溶解性有机碳含量,为Actinomycetota和Chloroflexota提供了多样化的代谢底物,Chloroflexota中部分类群在厌氧条件下对顽固性有机质的分解能力可能因套种根系交错形成的微氧环境而被激活[30]。苜蓿和将军菊苣通过生物固氮与吸磷功能的互补,减少了单一养分的竞争压力,Verrucomicrobiota与Actinomycetota形成代谢互惠网络,进一步稳定其生态位优势[25]。
在细菌属水平,优势菌属Gp6、Gp4、Gp2、Spartobacteria_genera_incertae_sedis在MJG和MMG(根际土样)中相对丰度均高于非根际和单作土样,这可能与套种模式下土壤养分和酶活性的协同提升有关。紫花苜蓿、甜高粱间作显著提高了根际有机质、有效磷含量及脲酶、蔗糖酶活性,而有机质与酶活性与变形菌门、酸杆菌门等优势菌群呈正相关,推测Gp类群可能通过参与碳磷代谢适应高有机质环境,此外,间作导致pH值降低,而酸敏感菌群(如酸杆菌门)的增殖可能间接促进相关功能菌的富集[13]。Gp3、Gp1在MJG土样中相对丰度低于MJCG土样,在MMG土样中高于MMCG土样,这可能和苜蓿与将军菊苣根际分泌物不同有关,苜蓿作为豆科植物分泌类黄酮等次生代谢物抑制Gp3和Gp1,而将军菊苣根系释放的糖类或酚酸能形成竞争性微环境[31],导致其根际Gp3、Gp1相对丰度较非根际减少,但相较于苜蓿非根际仍具优势。Gaiella在MJG和MMG(根际土样)中相对丰度均低于非根际土样,高于单作土样。这体现了木本与草本根系分泌物的协同调控作用,薄壳山核桃根系分泌酚酸类和萜类化合物,可直接抑制Gaiella的活性,导致其在根际土样中相对丰度低于非根际土样[32]。但套种将军菊苣和苜蓿通过减少单一化感物质的含量,减轻抑制作用,使Gaiella相对丰度高于单作土样[33]。
本研究发现,套种显著改变了真菌群落结构,Ascomycota在MJG土样中相对丰度最大,在MMG土样中相对丰度低于MMCG土样,但都显著高于单作土样,可能是套种模式通过改变根际土壤的养分有效性、酶活性及微生物互作,显著影响真菌群落的组成。郭童鑫等[34]对青稞、豌豆混作的研究发现,Ascomycota作为优势菌群(相对丰度63.5%~78.5%),其丰度与速效氮、有效磷和有机质等土壤因子密切相关。将军菊苣可通过根系分泌物或养分竞争增强根际土壤的酸化或有机质积累,从而促进Ascomycota在根际的富集。玉米与大蒜套作中,套作大蒜的根际土壤酶活性和微生物多样性显著提高,且真菌群落结构受土壤pH和养分有效性的影响[35]。苜蓿可通过固氮作用改变根际氮素形态,或通过根系分泌物(如酚酸类物质)抑制根际子囊菌的生长,导致其根际相对丰度低于非根际,这与甘蔗套种马铃薯的研究结论类似[36]。单作模式下,土壤微生物多样性通常较低,而间作通过种间互作增强根际效应,玉米和籽粒苋间作显著改变真菌群落结构,Ascomycota在单作玉米的土壤菌群中占比较低[37]。本研究发现,Mortierellomycota在MJG和MMG土样中相对丰度均显著高于非根际和单作土样,可能是将军菊苣和苜蓿的根系分泌物(如糖类、有机酸)能为Mortierellomycota提供偏好碳源,刺激其增殖[38],套种模式通过将军菊苣的深根碳和苜蓿的固氮作用优化土壤碳氮比,促进Mortierellomycota对复杂有机物的分解[39]。Glomeromycota、Paraglomeromycetes存在于套种土壤中,但在单作土壤中不存在;单作土壤中存在Tuber,而套种土壤中不存在。套种模式通过增加植物多样性,刺激丛枝菌根真菌和非典型菌根真菌的共生需求,而单作模式因宿主单一,菌根真菌多样性受限[40]。Tuber作为外生菌根真菌,偏好低氮环境,高氮条件可能抑制其孢子萌发或菌丝扩展[41]。
-
细菌和真菌作为土壤生态系统的重要组成部分,对土壤环境因子的改变比较敏感,其多样性和群落结构取决于细菌和真菌对土壤环境因子的适应能力[42],因此土壤微生物的多样性经常作为预测环境因子变化的指标[43]。对土壤理化性质与微生物细菌属、真菌属相关性的研究发现,土壤有效磷是细菌群落结构构建的主导因子,对将军菊苣套种薄壳山核桃的根际土壤细菌群落有促进作用,对单作及苜蓿套种土壤细菌群落有抑制作用,表明有效磷可通过改变根际pH值、酶活性及碳源分配,导致不同套种模式土壤细菌群落的差异[44]。将军菊苣根系分泌物富集溶磷菌,而苜蓿套种可能通过固氮作用改变氮磷比,抑制部分菌群[45]。AP、URE与Gp7,APH与Nitrospira呈显著正相关;SUC与Gaiella呈显著负相关。表明高磷可促进碳利用效率高的菌群,抑制依赖低磷寡营养策略的Gaiella菌属[46];APH与Nitrospira呈显著正相关,说明磷矿化能促进硝化作用[47]。
本研究发现,蔗糖酶和有效磷是真菌群落结构构建的主导因子,对单作及套种土壤的真菌群落存在抑制作用。在磷水平较低时,植物吸收磷素的主要方式是通过菌根吸收土壤中的多聚磷盐[48],Russula通过菌根共生协助植物磷吸收,二者在碳磷协同循环中形成互利关系,陈婷等[49]研究证实了菌根真菌与土壤磷元素间存在显著关系。
-
薄壳山核桃套种苜蓿和将军菊苣根际和非根际土壤细菌和真菌的α多样性差异无统计学意义,但根际土壤α多样性指数均高于非根际土壤。不同套种模式下细菌群落结构差异不明显,真菌群落结构差异有统计学意义。土壤有效磷是土壤细菌和真菌群落组成的共同主导环境因子。在生产中,通过增施磷肥提高土壤有效磷含量,能够促进植物对磷元素的吸收和利用。