


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
福白菊主要分布在湖北省麻城市北部山区,具有养肝明目、解毒消肿、散风清热等功效[1]。福白菊具有独特的种质资源和高品质特性,富含黄酮和绿原酸,是药食兼用型中药材[2-3],为湖北省重要道地药材。黄酮类化合物又称生物类黄酮,在人体中发挥着多种积极作用,如抗菌、消炎、降低血压、抗肿瘤及调节免疫功能等[4]。莲藕富含丰富的营养物质,具有很高的食用价值,然而,切分处理造成的机械损伤使莲藕细胞结构遭到破坏,容易发生褐变,导致感官品质和营养价值下降,货架期缩短,在一定程度上阻碍了鲜切莲藕的市场化进程[5-6]。传统保鲜剂抗坏血酸、柠檬酸、氯化钙都对鲜切果蔬的保鲜起重要作用,是作为保鲜剂的良好选择,但存在安全性或功能性缺陷,需开发天然成分替代方案,福白菊黄酮的强抗氧化特性为此提供了可能。福白菊和莲藕均为湖北独特资源的植物,福白菊黄酮复合保鲜剂不仅拓宽了福白菊的应用领域,也为莲藕储藏保存提供新思路,为湖北农业发展提供参考。
代谢组学运用高效的化学分析技术对生物体内所有小分子代谢物进行系统的鉴定与定量分析[7]。其中非靶向代谢组学作为一种全面的分析方法,有助于更深入地理解生物体的代谢过程[8]。近年来,已有莲藕褐变机理及酶变化的相关研究[9-10],然而,尚无利用非靶向代谢组学技术分析莲藕褐变进程中代谢物质的相关研究报道。本实验以氯化钙、抗坏血酸与柠檬酸作为复合保鲜剂,将是否添加福白菊黄酮提取物作为自变量,测定鲜切莲藕的理化指标,利用非靶向代谢组学技术对鲜切莲藕在褐变过程中的代谢产物进行研究,以阐明福白菊黄酮复合保鲜剂对延缓鲜切莲藕褐变的影响,并寻找发挥关键作用的代谢产物,为完善福白菊黄酮复合保鲜剂延缓鲜切莲藕褐变机制提供了理论参考。
全文HTML
-
莲藕购于荆门市掇刀区农贸市场;PE薄膜袋(长、宽为200 mm×200 mm,厚度为0.05 mm),广州健阳生物科技有限公司;磷酸二氢钠、磷酸氢二钠、硼酸、四硼酸钠、邻苯二酚、愈创木酚:分析纯,天津市华东试剂厂;抗坏血酸、氯化钙、柠檬酸,成都科隆化学品有限公司。
D3024R低温离心机,美国Scilogex公司;waters ACQUITY UPLC I-Class系统,美国沃特世公司;Hypesil Gold col-umn色谱柱(100 mm×2.1 mm,1.9 μm),美国Ther-mo Fisher公司。
-
挑选朵大、无虫害的优质福白菊花朵,将其置于40 ℃烘箱干燥至恒重,用超微粉碎机打碎成粉末状。将福白菊粉末与体积比70%的甲醇混合,料液质量体积比1∶49,超声温度58 ℃,70 min甲醇超声辅助提取后,于40 ℃低温旋转蒸发至不再有甲醇蒸出,再于烘箱40 ℃低温烘干成粉末状,迅速收集样品,避光保存于干燥器内。
以莲藕质量2 kg为参考标准,精确称取质量比0.15%福白菊黄酮提取物、0.5%氯化钙[11]、0.3%柠檬酸[12]、0.5%抗坏血酸[13]溶于2 L去离子水中,复配成保鲜剂备用。莲藕购买后迅速运至实验室,置于4 ℃冰箱中预冷6 h。预冷后仔细挑选无损伤、无腐烂变质且大小均匀的莲藕作为实验样品。将莲藕表皮的淤泥用清水清洗干净,再用已经消毒的不锈钢刀对莲藕进行断节切分以及去皮处理,将莲藕切成3mm左右的薄片,立即分别放入福白菊黄酮复合保鲜剂水溶液及不含福白菊黄酮的保鲜剂水溶液中浸泡30 min然后捞出,晾干水分。将经过福白菊黄酮复合保鲜剂处理过的鲜切莲藕称为样品实验组,经过不含福白菊黄酮的保鲜剂处理的鲜切莲藕称为对照组。每100 g用保鲜袋密封包装,于4 ℃冰箱冷藏。在0 d、6 d、12 d 3个时间对样品取样,测定其褐变度、多酚氧化酶(PPO)活性、过氧化物酶(POD)活性以及代谢产物。每个处理设3个重复。
-
测定鲜切莲藕的褐变度。称取2 g鲜切莲藕,放置于研钵中研磨,倒入离心管中,加入3 mL体积比95%的乙醇,放置于4 ℃下预冷6 h,在4 ℃下以10 000 r/min离心20 min,收集上清液,在波长410 nm下测量吸光值A[14],褐变度以10×A410表示。
-
采用分光光度法,取样品5 g,加入20 mL磷酸缓冲液,冰浴研磨后用纱布过滤,滤液在4 ℃下以8 000 r/min离心15 min,收集上清液,测前稀释适当倍数,备用。酶活反应体系包括5 mL邻苯二酚溶液和0.4 mL酶液,混合放置。反应体系在420 nm波长下有最大吸光值,吸光值在3 min内变化迅速,3 min后随着底物邻苯二酚被消耗,吸光值变化速度减缓。因此用在420 nm波长下3 min内吸光值的变化来表示PPO活性,每毫升酶液每分钟引起吸光值变化0.001为1个酶活单位[15-16]。
-
采用分光光度法,取样品5 g,加入20 mL磷酸缓冲液,冰浴研磨后用纱布过滤,滤液在4 ℃下以8 000 r/min离心15 min,收集上清液,测前稀释适当倍数,备用。酶活反应体系包括2 mL磷酸缓冲液、2 mL质量比2%的过氧化氢溶液、1 mL愈创木酚溶液、0.6 mL酶液,混合放置。反应体系在470 nm波长下有最大吸光值,吸光值在2 min内变化迅速,2 min后随着底物被消耗,吸光值变化速度减缓。因此用在470 nm波长下2 min内吸光值的变化来表示POD活性,每毫升酶液每分钟引起吸光值变化0.001为1个酶活单位。
-
参考文献[17]的方法,取用100 mg的莲藕样本,迅速使用液氮进行冷冻处理,并将其研磨至粉末状。接着,加入500 μL体积比80%的甲醇溶液,并利用振荡器充分混合。将含有莲藕代谢物的提取液置于冰上静置5 min,然后在4 ℃的环境下,以15 000 g的速度离心20 min,以获取上清液。随后,使用质谱级别的水将甲醇的体积比调整至53%。再次在4 ℃、15 000 g条件下离心20 min,从中取出100 μL的上清液。将样品通过0.22 μm的有机滤膜过滤,并将过滤后的样品转移到LC进样瓶中。在每个提取液中取30 μL,混合均匀,作为质控QC样本。使用53%的甲醇溶液作为空白对照,并按照与试验样品相同的步骤进行处理。将所有提取液进行标记并妥善保存于超低温冰箱中,以备后续进行质谱分析。
-
柱温:40 ℃;流速:0.2 mL/min;正离子模式:流动相A质量比0.1%甲酸,流动相B甲醇;负离子模式:流动相A 5 mmol/L醋酸铵(pH 9.0),流动相B甲醇;进样体积:10.0 μL;按照体积比进行梯度洗脱,洗脱程序为:[0,1.5) min,2%B;[1.5,14) min,2%到100%B;[14,17] min,100%到2%B。
质谱条件正负离子模式扫描范围100~1 500 m/z;质谱电压3 200 V,离子源温度320 ℃,GSⅠ和GSⅡ分别为276 000 Pa和69 000 Pa,采用数据依赖型串联质谱法进行MS/MS二级扫描[18]。
-
使用Compound Discoverer 3.1软件对莲藕样本的原始数据文件(.raw)进行处理。首先根据代谢物的保留时间、分子质量和质荷比等特征进行初步筛选。接下来,通过比较不同莲藕样本间的质量偏差和保留时间偏差来进行色谱峰的对齐。然后,将筛选和对齐后的数据与北京诺禾致源科技股份有限公司建立的代谢物数据库、mzCloud(
https://www.mzcloud.org/ )、mzVault等数据库中的信息进行比对,以鉴定莲藕中的代谢物种类。鉴定后,对莲藕样品中的每个色谱峰进行积分处理,每个色谱峰代表一种特定的代谢物,其相对含量用峰面积表示。 -
对莲藕进行不同储藏天数处理,将样本送到北京诺禾致源科技股份有限公司进行数据测序。本研究中,设置6种实验处理小组[实验组(0 d、6 d、12 d)、对照组(0 d、6 d、12 d)],每组测定时将6个平行样本混合成2个生物学重复,最终获得12个样本的代谢组测序数据集。
通过Compound Discoverer 3.1软件对莲藕在褐变过程中鉴定出的代谢物数据进行分析,实现代谢物的相对鉴定和定量。利用主成分分析(PCA)和正交偏最小二乘判别分析(OPLS-DA)等多变量统计方法来识别和分析莲藕样本中的显著差异代谢物。OPLS-DA方法用于评估模型的稳定性与准确性。利用多维统计VIP值、差异倍数(FC)以及q值分析莲藕褐变过程中的显著差异代谢物,显著差异代谢物满足FC大于等于1.200 0或小于等于0.833 3,且q<0.05两个条件。使用Heml 1.0程序对这些显著差异代谢物进行聚类分析,并根据分析结果绘制热图,以直观展示代谢物的变化情况。借助HMDB数据库(
https://hmdb.ca/metabolites )进行通路分析,从而揭示莲藕褐变过程中的代谢途径和机制。
1.1. 实验材料与仪器
1.2. 实验方法
1.2.1. 实验材料处理
1.2.2. 褐变度的测定
1.2.3. PPO活性测定
1.2.4. POD活性测定
1.2.5. 莲藕代谢物的提取
1.2.6. 色谱质谱条件
1.2.7. 代谢物定性与定量
1.3. 数据处理及分析
-
褐变是果蔬中酚类物质被氧化成褐色物质的表观体现[19],褐色物质的大小能够反映鲜切莲藕在储藏过程中的褐变程度。由表 1可知,随着储藏天数增加,鲜切莲藕的褐变度逐渐增大,褐变度与储藏天数呈正相关。在储藏期间,样品实验组的褐变度均低于对照组。储藏0~6 d鲜切莲藕褐变度的上升趋势高于6~12 d的上升趋势,可能是因为莲藕在鲜切加工和储藏过程中,因酚类物质与氧气接触发生酶促褐变。表明添加了福白菊黄酮提取物的保鲜剂更有效延缓鲜切莲藕的褐变。
-
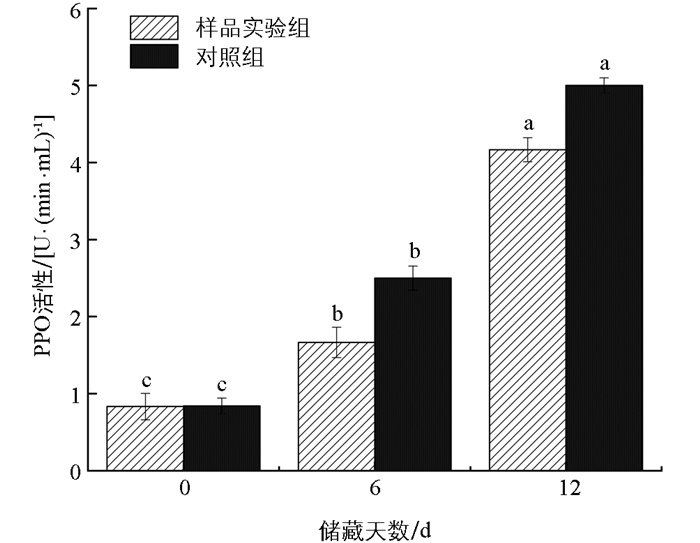
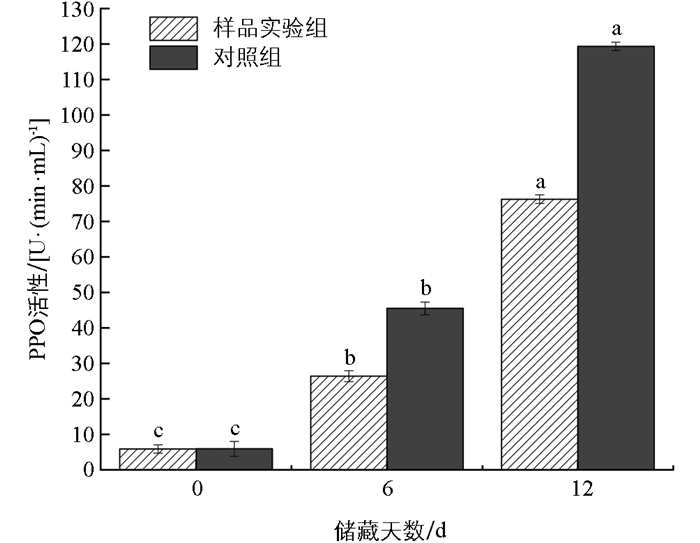
莲藕鲜切后由于机械损伤的存在,在储藏过程中较未鲜切的更易发生褐变,PPO和POD是褐变反应中的主要酶类[20]。PPO通常是引起果实褐变的一种重要酶类,不但影响果实口味还影响美观[21]。PPO是植物体内重要的呼吸酶类,其活性与果实的衰老程度相关[22]。在储藏期间,PPO活性逐渐升高,这可能是由于果实酶促反应其主要作用的酶是游离态多酚氧化酶(FPPO),切割破坏了细胞的膜结构,使得结合态多酚氧化酶(BPPO)从膜上游离成FPPO,与底物结合使其氧化,从而使PPO活性增高[23]。由图 1和图 2可知,在储藏初期,样品实验组和对照组的PPO活性和POD活性的差异无统计学意义。随储藏天数增加,样品实验组的PPO和POD活性比对照组更低。研究发现抗坏血酸对薄荷PPO活性[24]和洋蓟PPO活性[25]也有明显的抑制作用,随后的研究证实了抗坏血酸对PPO活性具有竞争性抑制作用[26-27]。
-
基于北京诺禾致源科技股份有限公司自建代谢物数据库、mzCloud、mzVault等数据库,对莲藕褐变过程中的代谢物进行定性和定量分析。表 2显示,在0 d样品实验组与对照组中共检测出1 888种代谢化合物,共筛选出1 146种显著差异代谢物,相对含量显著上调、下调的代谢物分别为746种和400种;在6 d样品实验组与对照组中共检测出1 921种代谢化合物,共筛选出671种显著差异代谢物,相对含量显著上调、下调的代谢物分别为421种和250种;在12 d样品实验组与对照组中共检测出1 941种代谢化合物,共筛选出1 031种显著差异代谢物,相对含量显著上调、下调的代谢物分别为513种和518种。
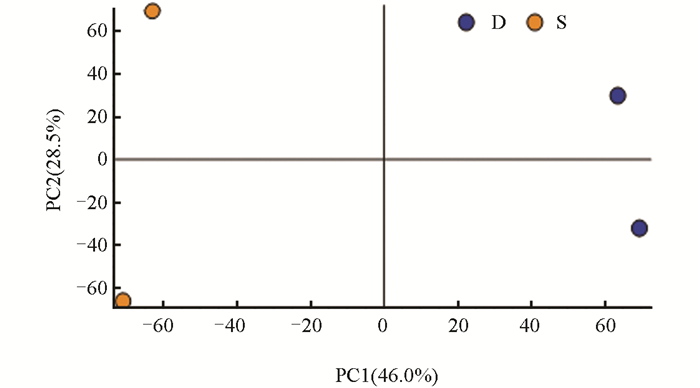
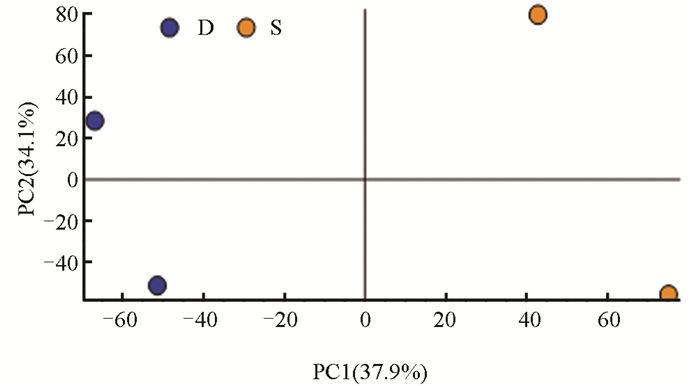
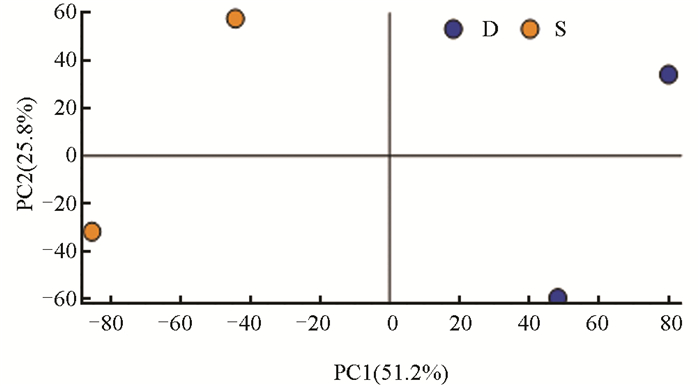
分别对样品实验组(S)和对照组(D)0 d、6 d、12 d(图 3、图 4、图 5)中的测序数据进行主成分(PCA)分析,以阐明本研究所使用的测序数据质量合格。
PCA结果显示,在0 d的样品实验组(S)和对照组(D)中,PC1、PC2的贡献率分别为46.0%,28.5%;在6 d的样品实验组(S)和对照组(D)中,PC1、PC2的贡献率分别为37.9%,34.1%;在12 d的样品实验组(S)和对照组(D)中,PC1、PC2的贡献率分别为51.2%,25.8%。结果显示,组间差异主要在第一主成分(PC1),组内差异主要在第二主成分(PC2),整体上看,组内差异远小于组间差异。每个样本在图上呈现出明显的分离趋势,提示每组中包含的代谢物表达量差异较大。为进一步研究代谢物差异情况,应结合差异代谢物筛选结果和聚类分析进行分析。
-
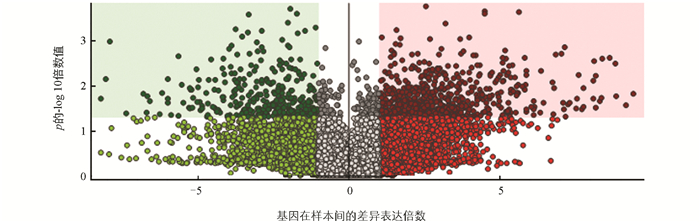
表 2显示,在0 d样品实验组和对照组中共检测出1 888种代谢化合物,共筛选出1 146种显著差异代谢物。其中有包括5-氟AKB48 N-(4-羟基戊基)代谢物(5-fluoro AKB48 N-(4-hydroxypentyl) metabolite)、DL-酪氨酸(DL-TYROSINE)、D-(+)-麦芽糖(D-(+)-Maltose)和美沙比妥(metharbital)等746种代谢物相对含量显著上调,占显著差异代谢物数的65.1%;有包括D-葡萄糖吡喃醛酸(D-glucopyranosiduronic acid)、棕榈酸(Palmitic Acid)和苯乙烯(Styrene)等400种代谢物的相对含量显著下调,仅为显著差异代谢物数的34.9%。有研究表明苯乙烯代谢物具有加速果肉褐变的作用[28],D-(+)-麦芽糖对碳水化合物活性酶的表达有抑制作用[29],并且在食品中,麦芽糖可以起到抑制果肉氧化的作用。这是因为麦芽糖具有一定的抗氧化性能,能够与自由基发生反应,从而减缓或阻止果肉的氧化过程。0 d样品实验组与对照组相比,苯乙烯的相对含量是显著下调的,可能是对莲藕添加福白菊黄酮提取物时,该代谢物被抑制,有助于延缓莲藕褐变;相反,D-(+)-麦芽糖代谢物的相对含量为显著上调,说明在添加福白菊黄酮后,莲藕中的碳水化合物降解过程减缓,从而延缓莲藕褐变。在0 d样品实验组与对照组代谢产物中检测出的主要代谢物之一是DL-酪氨酸,酪氨酸酶(tyrosinase)存在于CAT,与PPO、POD在半胱氨酸和侧链氨基酸的其他10个位置具有同源性,其部分保守结构域对这2种酶的功能有重要作用[30],有研究表明滇牡丹雄蕊提取物中的酪氨酸酶化合物在抗氧化和抑制PPO活性中起关键作用[31]。本研究为了更好展示样品实验组和对照组中显著差异代谢物表达情况,使用火山图作为可视化结果,如图 6所示。
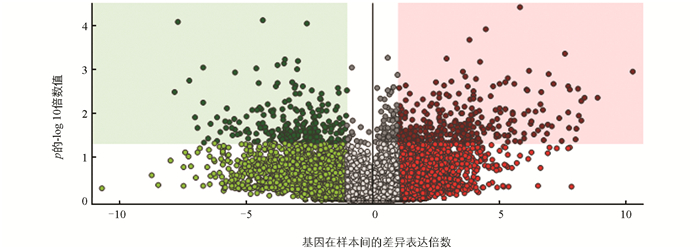
在6 d样品实验组和对照组中共检测出1 921种代谢化合物,共筛选出671种显著差异代谢物,相对含量显著上调、下调的代谢物分别为421种和250种。相对含量显著上调代谢物占显著差异代谢物数的62.7%,包括环唑嗪(Cyclazocine)、萘普生(Naproxen)和(+)-ar-姜黄酮((+)-ar-Turmerone)等;相对含量显著下调代谢物占显著差异代谢物数的37.3%,包括盐酸替来他明(tiletamine)、十一烯醛(undecenal)、七碳N-酰基-L-丙氨酸(C7-HSL)和4-[1, 2, 4]三唑并[4,3-b]吡啶并嗪-6-基硫代吗啉(4-[1, 2, 4]triazolo[4,3-b]pyridazin-6-ylthiomorpholine)。有研究表明(+)-ar-姜黄酮代谢物是潜在的食品抗真菌剂,防止黄曲霉在食品储存过程中生长、积累[32],七碳N-酰基-L-丙氨酸是诱导细菌产生细胞通讯的信号分子,可促进细菌的滋生[33]。结果表明,对莲藕添加福白菊黄酮提取物能激活(+)-ar-姜黄酮代谢物表达上调,抑制七碳N-酰基-L-丙氨酸代谢物表达,能有效抑制霉菌的滋生,可延缓莲藕褐变。在6 d样品实验组与对照组代谢产物中检测出的主要代谢物之一是(+)-ar-姜黄酮。文献[34]采用薄层色谱-生物自显影法对挥发油中倍半萜类化合物的抗氧化活性进行了测定和比较,确定莪术烯、呋喃二烯、α-姜黄酮、β-姜黄酮和β-倍半水芹烯为主要的抗氧化活性成分。姜黄酮会从PPO活性位点螯合Cu2+,也可能含有抑制PPO活性的一小段肽,从而抑制PPO活性[35]。差异分析可视化结果如图 7所示。
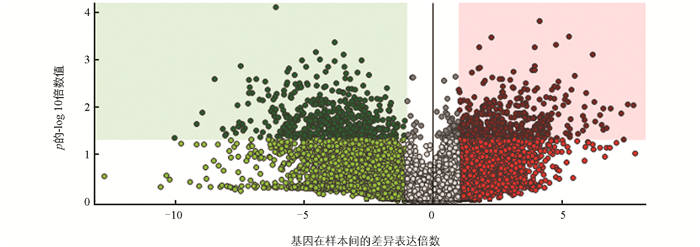
在12 d样品实验组和对照组中共检测出1 941种代谢化合物,共筛选出1 031种显著差异代谢物。有513种代谢物的相对含量显著上调,占显著差异代谢物数的49.8%,包括十五碳酸甲酯(Methyl 15-cyanopentadecanoate)、茉莉酮(Jasmonal)和己基肉桂醛(Hexyl cinnamaldehyde)等;有518种代谢物的相对含量显著下调,占显著差异代谢物数的50.2%,包括2-氨基-3-({[1-(二甲胺基)-1H-吡咯-2-基]甲亚胺基}亚)丁-2-二腈(2-amino-3-({[1-(dimethylamino)-1H-pyrrol-2-yl]methylidene}amino)but-2-enedinitrile)、L-同型半胱氨酸(L-Homocystine)、精胺(Spermine)和(E)-5-(甲基硫基)戊醛肟((E)-5-(methylsulfanyl)pentanal oxime)等。茉莉酮和异亮氨酸的结合物被证明是控制基因表达和次级代谢物产生的主要调节因子,防御事件的复杂级联是由相互作用的植物激素网络启动和控制的[36]。有研究表明,L-同型半胱氨酸会诱导细胞中的一氧化氮合酶依赖性脂质过氧化,过氧化会使得细胞损伤破裂[37]。在12 d样品实验组与对照组代谢产物中检测出的另一主要代谢物之一茉莉酮中茉莉酮酸甲酯可以增强果实中抗氧化酶包括SOD、POX、APX、MDHAR和DHAR的活性[38],这表明茉莉酮酸甲酯有增强水果抗氧化剂活性和清除自由基的能力[39]。结果显示,添加福白菊黄酮提取物有助于激活莲藕的防御机制,并且能有效抑制细胞过氧化,增加莲藕抗氧化能力,抑制莲藕褐变。差异分析可视化结果如图 8所示。
上述结果显示,随着实验天数的增加,实验组中相对含量上调代谢物占比减少,而相对含量下调代谢物占比增加。所以,添加福白菊黄酮提取物可激活鲜切莲藕的防御机制,抑制细胞过氧化以及细菌滋生,从而延缓莲藕褐变的发生。
-
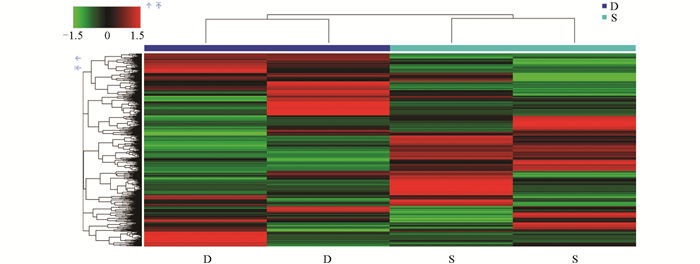
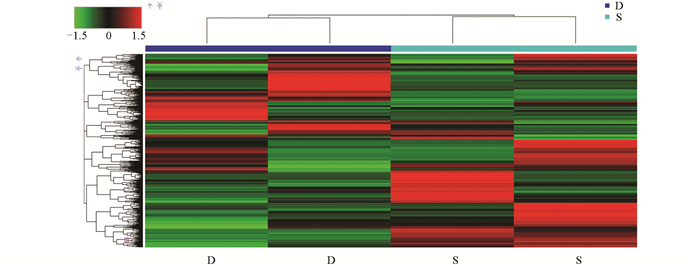
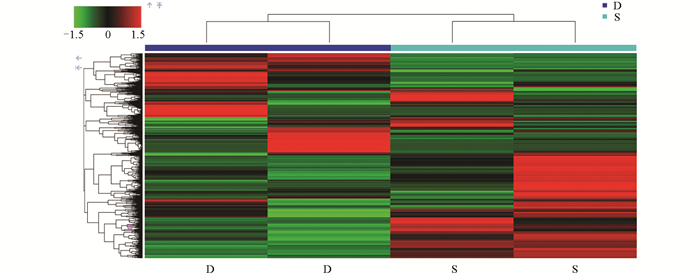
采用层次聚类(Hierarchical Clustering)方法对0 d、6 d、12 d的样品实验组(S)和对照组(D)进行差异代谢物表达量分析(可视化结果如图 9-图 11)。图中颜色越红表示莲藕中代谢物的相对含量越高,越绿则表示莲藕中代谢物的相对含量越低。结果显示,上述的苯乙烯代谢物在对照组0 d中呈红色,在样品实验组0 d中呈绿色,而D-(+)-麦芽糖的结果相反,符合预期。同样,(+)-ar-姜黄酮和茉莉酮代谢物分别在对照组6 d和12 d中呈绿色,在样品实验组6 d和12 d中呈红色,而七碳N-酰基-L-丙氨酸和L-同型半胱氨酸结果相反,验证了上述差异代谢物分析结果。
2.1. 褐变度分析
2.2. PPO活性和POD活性测定
2.3. 差异代谢物测定
2.4. 差异代谢物分析
2.4.1. 差异代谢物筛选结果分析
2.4.2. 差异代谢物层次聚类分析
-
在储藏期间,样品实验组的褐变度明显低于对照组。添加了福白菊黄酮提取物的保鲜剂可以抑制PPO和POD的活性,延缓鲜切莲藕的褐变。通过对样品实验组和对照组进行代谢物分析,发现在不同实验天数的样品中存在大量显著差异代谢物。在0 d样品实验组与对照组中筛选出1 146种显著差异代谢物,在6 d样品实验组与对照组中筛选出671种显著差异代谢物,在12 d样品实验组与对照组中筛选出1 031种显著差异代谢物。其中DL-酪氨酸、(+)-ar-姜黄酮、茉莉酮对PPO和POD的活性呈抑制作用。在具体的代谢物分析中,一些代谢物的含量在样品实验组和对照组之间呈现显著差异,例如苯乙烯、(+)-ar-姜黄酮和茉莉酮等代谢物的表达在不同条件下呈现出相反的趋势,这进一步验证了添加福白菊黄酮提取物对莲藕代谢物表达的影响。
综上所述,添加福白菊黄酮提取物通过激活防御机制和调节代谢物表达可对延缓鲜切莲藕褐变起到一定作用,研究结果为莲藕保鲜及莲藕褐变机理提供了理论依据。