



 下载:
下载:

-
开放科学(资源服务)标志码(OSID):

-
生物胺是无脊椎动物体内具有重要生物活性的小分子化合物,主要包括章鱼胺(octopamine,OA)、酪胺(tyramine,TA)、组胺(histamine,HA)、多巴胺(dopamine,DA)和5-羟色胺(serotonin,5-HT)[1]. 章鱼胺作为昆虫体内不可或缺的一类生物胺,参与昆虫的昼夜节律、内分泌、好斗、飞行以及学习与记忆等重要生理活动[2]. 此外,还有研究证实,章鱼胺在应对不同环境胁迫时也发挥着积极的功能.
章鱼胺的功能行使需特异性地结合章鱼胺受体[3](octopamine receptor,OARs). 章鱼胺受体是典型的G蛋白偶联受体(G-protein coupled receptor,GPCR). 自第一个章鱼胺受体基因从黑腹果蝇(Drosophila melanogaster)中成功克隆以来[4],目前已在烟草天蛾(Manduca sexta)、家蚕(Bombyx mori)、赤拟谷盗(Tribolium castaneum)、棉铃虫(Helicoverpa armigera)等多种昆虫中发现[5-8]. 随着基因组学的快速发展,章鱼胺受体基因研究更加系统全面. 章鱼胺受体除了与OA结合发挥重要生理功能外,还是多种杀虫(螨)剂的作用靶标,如杀虫剂杀虫脒和杀螨剂双甲脒[9]. 因此,针对昆虫、螨类章鱼胺受体特异性开发激动剂和拮抗剂,可以创制高效、安全的新型杀虫剂.
巴氏新小绥螨(Neoseiulus barkeri),隶属于蛛形纲(Arachnida)蜱螨亚纲(Acari)寄型螨目(Parasitiformes)植绥螨科(Phytoseiidae)新小绥螨属(Neoseiulus)[10]. 其分布范围广、捕食能力强、易规模化生产,是目前应用最为广泛的生物防治天敌之一[11],对叶螨、蚜虫、粉虱等害虫(螨)具有理想的控制作用[12]. 研究巴氏新小绥螨章鱼胺受体基因,可以针对性地研发对其安全的新型杀虫(螨)剂,而目前有关其章鱼胺受体基因的研究还未见报道. 鉴于此,本研究鉴定了巴氏新小绥螨6个章鱼胺受体基因,分析巴氏新小绥螨章鱼胺受体基因不同发育阶段、不同温度及饥饿胁迫下的表达模式,为进一步深入研究章鱼胺受体的生理功能和药理学提供理论依据.
全文HTML
-
供试螨源:巴氏新小绥螨为西南大学柑桔研究所实验室多年继代饲养种群. 以椭圆食粉螨(Aleuroglyphus ovatus)进行饲喂,并于人工气候箱(温度25±1 ℃,相对湿度75%~85%,光周期14L/10D)中饲养.
-
挑取足量的巴氏新小绥螨雌成螨于洁净的凹槽玻璃板中饲养,任其产卵. 不同发育阶段螨源和不同温度处理后螨源定期饲喂足量椭圆食粉螨,饥饿处理螨源的收集则待卵孵化至成螨后停止饲喂椭圆食粉螨.
不同发育阶段螨源的收集:分别收集巴氏新小绥螨800粒卵,500头幼螨,500头若螨,200头雌成螨,重复3次.
不同温度处理后雌成螨和幼螨的收集:将光照培养箱的温度分别设置为15 ℃,20 ℃,25 ℃,30 ℃和35 ℃,24 h后分别收集巴氏新小绥螨幼螨500头,雌成螨200头,重复3次.
饥饿处理后雌成螨的收集:将饥饿处理时间分别设置为0 h,6 h,12 h,18 h和24 h,每个处理分别收集巴氏新小绥螨雌成螨200头.
使用ReliaPrepTM RNA Tissue Miniprep System (普洛麦格)试剂盒提取巴氏新小绥螨样本总RNA,利用紫外分光光度计和1%无核酶的琼脂糖凝胶电泳检测RNA的质量和浓度. 将质量合格的mRNA样本按照反转录试剂盒HiScript Ⅲ 1st Strand cDNA Synthesis Kit (+ gDNA wiper) (诺唯赞)的步骤,合成cDNA模板.
-
从巴氏新小绥螨转录组数据(西南大学柑桔研究所实验室构建,NCBI数据库登录号:SRP333450)中筛选出注释为章鱼胺受体基因的序列,使用NCBI在线软件ORF Finder(
http://www.ncbi.nlm. nih.gov/gorf/gorf.html ) 对筛选的章鱼胺受体基因进行开放阅读框预测和蛋白质翻译,将翻译后的氨基酸序列用Smart BLAST来验证,验证为章鱼胺受体的基因再次进行BLASTX比对,确认候选章鱼胺受体基因的命名.利用Primer Premier 5.0软件设计PCR引物(表 1),引物由北京擎科生物技术有限公司进行合成. 以巴氏新小绥螨雌成螨cDNA为模板,利用高保真DNA聚合酶(诺唯赞)进行PCR扩增. PCR反应体系(50 μL):反转录产物2 μL,上游引物和下游引物各2 μL,酶25 μL,RNase-free dd H2O 19 μL. 反应程序:95 ℃ 30 s; 95 ℃ 15 s,72 ℃ 15 s,72 ℃延伸30 s,35个循环. 将目的片段通过DNA回收试剂盒(诺唯赞)纯化后连接至T/A Blunt Vector载体,然后转化至大肠杆菌Escherichia coli DH5α感受态细胞. 在含有卡那霉素的LB固体培养基上过夜培养,挑取阳性克隆于LB液体培养基中,37 ℃下180 r/min孵育3 h后进行菌液PCR鉴定,将验证正确的菌液送至北京擎科生物技术有限公司进行测序.
将测序验证的ORF序列利用TMHMM Server v. 2.0等在线软件进行跨膜结构域、分子量和等电点预测; 然后使用DNAMAN V6软件进行多重序列比对; 再应用MEGA 7.0软件中的邻接法构建系统发育树,1 000次检测; 最后应用DNAMAN V6软件对不同无脊椎动物中章鱼胺受体基因的氨基酸序列进行同源性比对.
-
提取样品的RNA并合成cDNA模板. 利用Primer Premier 5.0软件设计qRT-PCR引物(表 2),根据ChamQ Universal SYBR© qPCR Master Mix (诺唯赞)试剂盒进行qRT-PCR检测. qRT-PCR在qTOWER qPCR system (Analytik Jena)上进行,反应体系(20 μL):2 × ChamQ SYBR qPCR Master Mix 10 μL,上游引物和下游引物各0.25 μL,ChamQ Universal SYBR© qPCR Master Mix 5 μL,cDNA模板0.4 μL,RNase-free dd H2O 4.1 μL. 反应程序:95 ℃ 2 min; 95 ℃变性30 s,60 ℃退火延伸30 s,40个循环; 熔解曲线60 ℃ 30 s,95 ℃ 2 min. 选择泛素结合酶UBC(Ubiquitin Conjugating Enzyme,KP310123)基因作为内参基因[13],采用2-ΔΔCt法[14]计算基因的相对表达量. 每个样品设置3个生物学重复,每个生物学重复进行2次技术重复.
-
采用SPSS 26.0软件对实验数据进行统计分析,通过单因素方差分析法(one-way ANOVA)和Duncan氏新复极差法(p<0.05)对候选基因的相对表达量进行差异显著性分析,以x±s表示.
1.1. 供试生物材料
1.2. 不同螨态、胁迫条件的收集和保存处理方法
1.3. 巴氏新小绥螨章鱼胺受体基因的鉴定和生物信息学分析
1.4. 定量PCR
1.5. 数据分析
-
根据在线软件TMHMM Server v. 2.0分析,巴氏新小绥螨 NbOctα1R1 ,NbOctα1R2 ,NbOctTyrR ,NbOctβ2R1 ,NbOctβ3R (图 1)都具备7个跨膜结构域(TM1-TM7)、胞内C端、胞外N端、胞内环和胞外环各3个,位于TM5和TM6之间的第3个胞内环最长,这些都是G蛋白偶联受体的典型特征,表明上述基因的cDNA序列完整. NbOctβ2R2 拥有5个跨膜结构域,无TM1和TM2. 筛选鉴定获得的6个章鱼胺受体基因分别命名为 NbOctα1R1 (GenBank登录号:MZ855515),NbOctα1R2 (GenBank登录号:MZ855512),NbOctTyrR (GenBank登录号:MZ855514),NbOctβ2R1 (GenBank登录号:MZ855513),NbOctβ2R2 (GenBank登录号:MZ855511),NbOctβ3R (GenBank登录号:MZ855516). 由表 3可知,巴氏新小绥螨6个章鱼胺受体基因序列的完整开放阅读框(ORF)为936~1 983 bp,编码312~661个氨基酸,分子量为35.24~72.43 kD,等电点为8.47~9.63.
为确定上述6个章鱼胺受体基因的分类地位和进化关系,将其与已报道的17个无脊椎动物的55个OARs家族的氨基酸序列构建系统进化树(图 2). 结果表明,巴氏新小绥螨 NbOctα1R1 ,NbOctα1R2 ,NbOctTyrR 与西方静走螨(G. occidentalis) GoOctα1R1 同源性达86.57%~95.73%,与其他几种无脊椎动物同源基因的同源性在70.00%~90.00%之间; 同属肾上腺素能β受体的 NbOctβ2R1 ,NbOctβ2R2 ,NbOctβ3R 章鱼胺受体基因与西方静走螨、雅氏瓦螨(Varroa jacobsoni)、狄氏瓦螨(Varroa destructor)的章鱼胺受体基因同源性较高,均在80.00%以上.
-
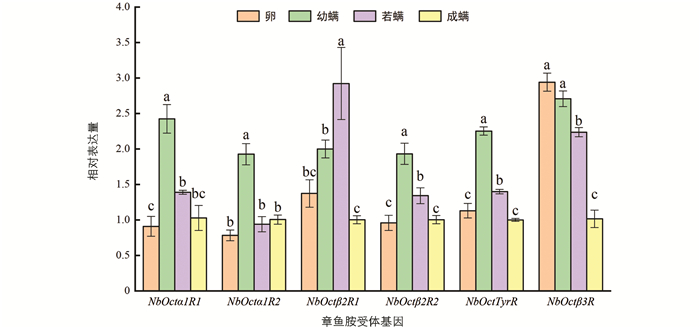
巴氏新小绥螨6个章鱼胺受体基因在卵、幼螨、若螨和成螨中都有所表达(图 3). 同属肾上腺素能α1受体的 NbOctα1R1 ,NbOctα1R2 在不同螨态中的表达模式与属于肾上腺素能α2受体的 NbOctTyrR 具有相似性,均在幼螨中表达量最高(p<0.05). 同属肾上腺素能β受体的 NbOctβ2R1 ,NbOctβ2R2 和 NbOctβ3R 在不同发育阶段的表达情况具有一定的差异,其中 NbOctβ2R1 在成螨中表达量最低,在若螨中表达量最高(p<0.05); NbOctβ2R2 在幼螨中表达量最高(p<0.05),在卵中表达量最低; NbOctβ3R 在卵和幼螨中表达量高(p<0.05).
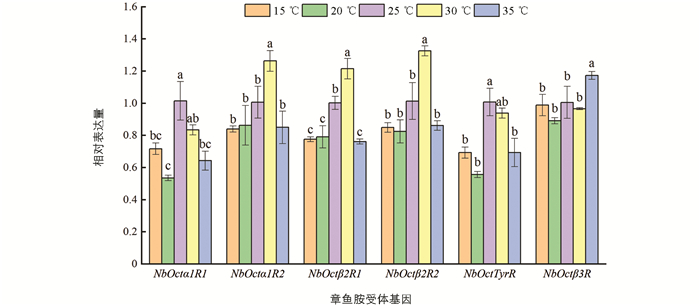
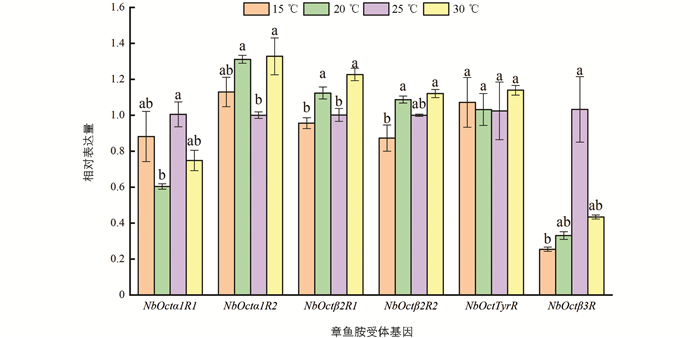
温度对巴氏新小绥螨成螨章鱼胺受体基因的影响如图 4. NbOctα1R2 ,NbOctβ2R1 ,NbOctβ2R2 的表达量随温度的升高而升高,30 ℃时的表达量最高; NbOctα1R1 ,NbOctTyrR 在25 ℃时的表达量最高,随后随温度的升高表达量下降; NbOctβ3R 在35 ℃时表达量显著高于其他温度下的表达量(p<0.05). 幼螨中(图 5)章鱼胺受体基因 NbOctβ2R1 ,NbOctβ2R2 在高温时表达量上调,低温时表达量下调; 在 NbOctTyrR 下的表达量均没有显著变化(p>0.05); NbOctβ3R 的表达量在低温时最低,在25 ℃时最高(p<0.05); 其他基因表达量在低温和高温下没有明显规律性.
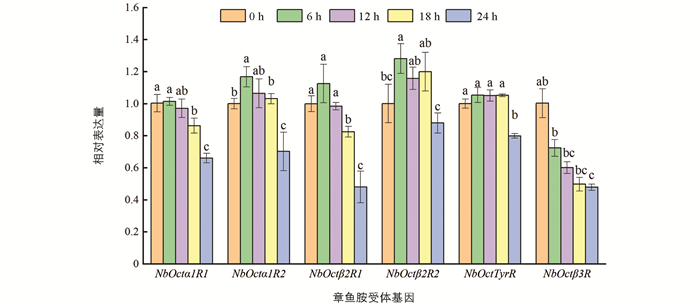
经过6 h,12 h,18 h和24 h的饥饿处理后,巴氏新小绥螨6个章鱼胺受体基因的表达情况如图 6,NbOctβ3R 表达量随饥饿时间增加而逐渐下降,其余章鱼胺受体基因表达量先有不同程度的升高,在24 h的表达量均显著低于0 h(p<0.05).
2.1. 巴氏新小绥螨章鱼胺受体基因的鉴定和生物信息学分析
2.2. 巴氏新小绥螨不同发育阶段、不同温度及饥饿胁迫下的表达模式分析
-
章鱼胺对特定器官或细胞的调控依赖于细胞膜上的章鱼胺受体传递神经信号. 随着研究的不断深入,越来越多的章鱼胺受体在不同的昆虫物种中得到鉴定,依据结构和信号传递方式将章鱼胺受体分为3类:肾上腺素能α受体(α-adrenergic-like octopamine receptors),肾上腺素能β受体(β-adrenergic-like octopamine receptors)和章鱼胺酪胺受体(octopamine/tyramine receptors)[15]. 本研究从巴氏新小绥螨(N. barkeri)转录组数据库中鉴定到6个章鱼胺受体基因,分别为 NbOctα1R1 ,NbOctα1R2 ,NbOctTyrR ,NbOctβ2R1 ,NbOctβ2R2 ,NbOctβ3R ,其中肾上腺素能β受体OARs家族有2类(NbOctβ2R 和 NbOctβ3R),未发现Octβ1R类受体,这与果蝇(Drosophila)[16]肾上腺素能β受体家族拥有Octβ1R,Octβ2R和Octβ3R共3类受体不同,同时意大利蜜蜂(Apismellifera ligustica)肾上腺素能β受体家族存在第4类受体基因 AmOctβ4R [17],这表明不同昆虫的章鱼胺受体存在种间特异性. 巴氏新小绥螨章鱼胺受体基因在进化关系上与西方静走螨最为接近,目前关于西方静走螨章鱼胺受体基因功能的研究较少. 在黑腹果蝇(D. melanogaster)[18]、二化螟(Chilo suppressalis)[19]、褐飞虱(Nilaparvata lugens)[20]的研究中,发现章鱼胺受体均存在两个剪切体,而在巴氏新小绥螨Octα1R类受体基因(NbOctα1R1 ,NbOctα1R2),Octβ2R受体基因(NbOctβ2R1 ,NbOctβ2R2)中不存在剪切体,可能是由于不同昆虫的章鱼胺受体的功能存在差异.
昆虫中不同基因在不同组织和发育阶段的表达量存在差异,还受多种外界因素的影响,这与其功能息息相关. 本研究结果表明巴氏新小绥螨6个章鱼胺受体基因在不同螨态均有表达,且在幼螨和若螨中表达水平较高,这与黑腹果蝇 DmOctβR1 在幼虫中高表达相似[21]. 其他昆虫也有相似结果,赤拟谷盗[7]、棉铃虫[8]、二化螟[22]体内 OctβR2 在不同发育阶段均有所表达,尤其在幼虫期高表达. 本研究结果表明巴氏新小绥螨章鱼胺受体基因 NbOctα1R1 ,NbOctα1R2 ,NbOctTyrR ,NbOctβ2R2 可能在幼螨阶段发挥着重要的作用,NbOctβ2R1 可能在若螨阶段发挥着重要的作用,这可能与不同发育阶段独特的生活习性相关. 除在幼螨和若螨中高表达外,NbOctβ3R 在卵中表达水平也较高,这与黑腹果蝇 DmOctβ2R 表达模式相似[23],推测 NbOctβ3R 可能具有诱导虫体产卵和促进卵发育的重要作用. 此外,温度、药剂胁迫等外界条件的改变会引起昆虫体内章鱼胺含量的改变,有研究发现赤拟谷盗章鱼胺的含量随着温度的变化发生改变[24]. 本研究结果表明成螨中 NbOctβ3R 受高温影响大,在35 ℃时表达量最高,其所调控的功能可能受到高温的影响而增强,而其他基因的表达量在15 ℃和35 ℃下均较低,表明高温和低温很可能会减弱基因调控的功能; 而在幼螨中,NbOctβ2R1 ,NbOctβ2R2 在15 ℃下的表达量显著低于35 ℃下的表达量(p<0.05),表明低温可能会抑制基因的表达,高温可能会增强基因的表达. 在对巴氏新小绥螨进行饥饿胁迫后发现,6个章鱼胺受体基因在24 h的表达量都最低,NbOctα1R2 在6 h的表达量显著高于0 h(p<0.05). 有研究发现果蝇在饥饿条件下,体内章鱼胺(OA)含量升高,机能亢进,虫体活动能力增强[25],黑腹果蝇中 DmOctβ2R 已被证明对昆虫运动行为具有重要的调控作用[26]. 由此推测饥饿胁迫6 h时引起巴氏新小绥螨OA水平升高,NbOctα1R2 表达量显著上调(p<0.05),活动能力增强,有利于寻觅猎物. 章鱼胺受体基因在幼螨和若螨阶段总体高表达,受温度和饥饿胁迫影响变化情况存在差异,可能是由于不同的章鱼胺受体基因功能存在差异以及响应外界变化的机制不一.
大量研究表明,章鱼胺受体在特定组织中有特异性功能,后被证实与视觉和嗅觉信息处理有关[27]. 黑腹果蝇输卵管上皮细胞表达的章鱼胺受体OAMB,参与调控输卵管中卵的运输[28]; 美洲大蠊(Periplaneta americana)触角中的 PaOA1则与嗅觉功能相关[29]. 然而巴氏新小绥螨体型较小,无法人工解剖分离其器官组织,这就限制了对巴氏新小绥螨不同组织中章鱼胺受体表达模式的研究. 本实验仅从转录水平上揭示不同章鱼胺受体基因间表达的差异性,未来还可根据巴氏新小绥螨章鱼胺受体的特异性开发激动剂和拮抗剂,并进行药理学分析,进而研制出可以高效防治害螨并对益螨安全的新型杀虫剂.