


 下载:
下载:
-
开放科学(资源服务)标志码(OSID):

-
喀斯特(岩溶)地区是全球最主要的生态脆弱带之一,占据了约12%的世界陆地总面积[1-2]. 喀斯特地区土壤资源极其匮乏,稀少的土壤与裸露的基岩镶嵌而布[3-4]. 西南喀斯特地区降雨酸度大,以碳酸盐为主要成分的成土母岩渗漏性强、不耐溶蚀[5],使得岩体上存在着大量岩溶裂隙[6],降雨后会导致表层土壤沿其流失. 因此,岩溶裂隙层厚度不同,裂隙土壤小生境中的水分、养分、氧气、空间等便也存在差异[7],使其在垂直方向表现出异质性[8]. 在全球气候变化的背景下,西南喀斯特地区呈现出降雨总量不变、全年雨日明显减少的趋势[9]. 单次降雨强度增大,土壤易被冲散,水土流失更为严重,加剧了土壤小生境的垂直异质性,进一步影响着植物的生长.
植物光合光响应曲线揭示了净光合速率与光合有效辐射之间的对应关系. 通过光合光响应曲线获得的生理参数(包括表观量子效率、最大净光合速率、光补偿点、光饱和点以及暗呼吸速率)[10-15],能直接或间接反映出植物的生理生态学过程. 目前,最常用的3种光合光响应数学模型分别是直角双曲线模型[16]、非直角双曲线模型[17]和指数模型[18],但通过这3种光响应模型求得的光饱和点低于实测值,而最大净光合速率大于实测值,且无法拟合光抑制情况下的光响应曲线[11-12]. 叶子飘等[12, 19]提出了1种直角双曲线修正模型,此模型可以克服上述3种模型的缺陷,但目前主要应用于农作物和草本植物的光合光响应的模拟,在木本植物方面应用较少[20].
桢楠(Phoebe zhennan)为樟科楠属的常绿乔木,是珍贵的用材树种,在中国被称为“金丝楠木”,具有非常重要的经济价值[21-23]. 由于历年来对桢楠的滥砍滥伐,其原生林几乎被毁,且由于桢楠后期生长缓慢和遗传多样性较低等原因,导致现存资源有限且分布零散[24]. 研究表明桢楠喜湿耐荫,适宜在山腰山谷中生存. 我们在野外调查中发现,重庆市大足区及永川区的喀斯特环境中均存在长势良好的桢楠林,而当地有关部门也正在喀斯特地区尝试推广桢楠的种植. 因此,本研究以桢楠幼苗为研究对象,探讨不同降雨时间格局下,喀斯特垂直异质生境中桢楠幼苗的光合光响应曲线的拟合及光响应参数特征,以期为桢楠在喀斯特地区的种植及保护提供参考.
全文HTML
-
供试植物为1年生桢楠幼苗,供试土壤与岩石分别为黄色石灰土与喀斯特石灰石,二者均取自具有典型喀斯特地貌的重庆沙坪坝区中梁山. 黄色石灰土的理化性质:pH值为7.4±0.14,有机质0.34%±0.02%,全氮0.28±0.03 g/kg,全磷0.39±0.02 g/kg,全钾23.7±3.22 g/kg,田间持水量39.80%±2.23%.
-
采用双因素随机区组实验,共设置垂直小生境和降雨时间格局两个处理. 垂直小生境处理分为无石全土(S0)、半石半土(S1/2)和多石少土(S3/4). 为模拟这3种水平,自制容器以保证各生境总体积相同(54 cm×39 cm×24 cm). S0生境为全土;S1/2生境上半部分设置土壤层,下半部分设置岩溶裂隙层;S3/4生境上1/4为土壤层,下3/4为岩溶裂隙层. 依据重庆市年平均最长连续无有效降水日数为19 d[25],将降雨时间格局处理分为2 d降雨间隔(I2d)和19 d降雨间隔(I19d). 为模拟这2种水平,根据重庆市沙坪坝区日平均降雨量及容器底面积大小,将I2d的4个季度单次施水量分别设置为600,3 000,2 900,1 100 mL,将I19d设置为4 000,20 300,19 500,7 600 mL,两种水平的总施水量相同. 实验在重庆市北碚区西南大学生态园实验基地(29°49′N,106°25′E)大棚内进行,海拔高度249 m,大棚顶部透明,周围开放. 桢楠幼苗于2019年7月15日移栽,8月14日进行水分处理. 实验共设置18个重复.
-
处理1年8个月后,在每一处理中选取3株长势一致的幼苗,于2021年3月23-25日使用Li-6400便携式光合仪(美国Li-COR公司)自上午8:30起测定其光响应曲线. 测定时采用内置红蓝光源(6400-02B Red Blue Light Source,Li-COR),气体流速设置为500 μmol/s,光合有效辐射(PAR,以I表示)从高到低依次设置为1 500,1 200,1 000,800,600,400,200,150,100,50,20,0 μmol/(m2·s),选用自动测量模式进行测量,每一设定值下停留120~200 s. 每天测定一组重复中的6株,每株选取上部受光方向一致、位置相近并且成熟健康的1片叶子进行测量.
-
分别选用直角双曲线模型(RHM)、非直角双曲线模型(NRHM)、直角双曲线修正模型(MRHM)和指数模型(EM)拟合桢楠幼苗的光响应曲线,各模型的计算公式[16-18, 26]分别为
直角双曲线模型:
非直角双曲线模型:
直角双曲线修正模型:
指数模型:
式中,Pn表示净光合速率,I表示光合有效辐射,α表示初始量子效率,Pnmax表示最大净光合速率,Rd表示暗呼吸速率,θ表示非直角双曲线模型中曲线的弯曲程度;β,γ是直角双曲线修正模型中的2个修正系数,β表示光抑制项,γ表示光饱和项,e表示自然常数. 实验数据使用光合计算软件(4.1.1版)处理,用各模型拟合光响应曲线及最大净光合速率(Pnmax)、光补偿点(IC)、暗呼吸速率(Rd)这些光响应参数,通过求算光合有效辐射I≤200 μmol/(m2·s)下光响应曲线的直线拟合方程的斜率[27],得到表观量子效率(AQE,以EAQ表示).
为了对光响应曲线拟合模型的拟合效果进行定量的评估,本研究引入了决定系数(R2)、均方根误差(ERMS)和平均绝对误差(EMA)来检验各模型的拟合优度[28-30]. R2越接近1,ERMS和EMA越小,拟合精度越高. 使用光合计算软件(4.1.1版)直接得出R2,使用如下计算公式得出ERMS和EMA:
式中,n表示实测值个数,Yo,i表示净光合速率的实测值,Ym,i表示净光合速率的模型拟合值.
-
用拟合优度最高的模型拟合各处理中3个重复的光响应曲线,分别计算其最大净光合速率、光补偿点、暗呼吸速率及表观量子效率. 使用SPSS 26.0对相同降雨时间格局下不同小生境中桢楠幼苗的光响应参数进行单因素方差分析(One-Way ANOVA);对相同小生境中不同降雨时间格局下桢楠幼苗的光响应参数进行t检验;对不同降雨时间格局下不同小生境中桢楠幼苗的光响应参数进行双因素方差分析(Two-Way ANOVA),用Origin 2021作图.
1.1. 实验材料
1.2. 实验方法
1.3. 光响应曲线测定
1.4. 光响应曲线拟合
1.5. 数据处理
-
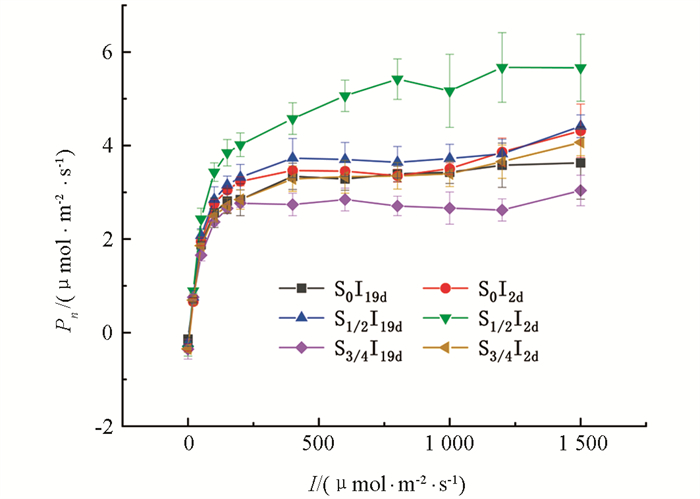
当光合有效辐射I≤600 μmol/(m2·s)时,随I增加,各处理下桢楠幼苗的净光合速率(Pn)均不断升高,当I达到约600 μmol/(m2·s)后,低频降雨下多石少土(S3/4I19d)这一处理中的桢楠幼苗开始出现光抑制现象,其Pn随I的增加而降低,其他各处理的Pn仍随I的增加而升高. 在相同光合有效辐射时,高频降雨下半石半土(S1/2I2d)这一处理中的桢楠幼苗的Pn最大,低频降雨下多石少土(S3/4I19d)这一处理中的桢楠幼苗的Pn最小,其他4种处理(S0I19d,S0I2d,S1/2I19d,S3/4I2d)的Pn相差不大. S0I2d,S1/2I19d,S3/4I19d,S3/4I2d在I > 1 200 μmol/(m2·s)时Pn还有所增加,可能是在所设定的测量光有效辐射范围内尚没有光饱和现象出现(图 1).
-
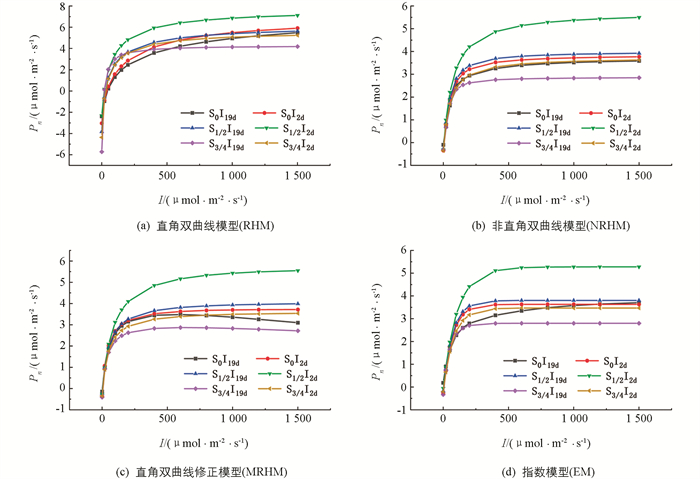
直角双曲线模型(RHM)、非直角双曲线模型(NRHM)、直角双曲线修正模型(MRHM)和指数模型(EM)对桢楠光响应过程的拟合结果见表 1和图 2. 结果表明,无论是哪种处理,由NRHM拟合出的光响应曲线的R2都是最大的,ERMS和EMA相对较小,说明NRHM的拟合精度最高;MRHM,EM的拟合精度也较高,R2均在0.87以上,多数ERMS和EMA略高于NRHM;相比于其他3种模型,RHM的R2最小,其最大值只有0.762 3,ERMS和EMA较大,拟合精度较差(表 1).
然而,在评估各光响应模型的拟合优度时,不能仅仅考虑其拟合精度,还要比较其拟合效果. 由图 2可知,由实测值绘得的光响应曲线并非一直呈现上升趋势,在光合有效辐射达到一定程度后,净光合速率开始随着光合有效辐射的增加而下降,即表现出光抑制现象. 但RHM,NRHM,EM 3种模型在达到光饱和之后的拟合点均高于实测值,拟合曲线无限上升,无法拟合出光抑制现象,只有MRHM具有最高的拟合优度.
-
因直角双曲线修正模型(MRHM)拟合优度最佳,故使用其来分析拟合降雨时间格局下喀斯特垂直异质生境中桢楠幼苗的光响应曲线,得出各处理的光响应参数并作统计学检验分析.
-
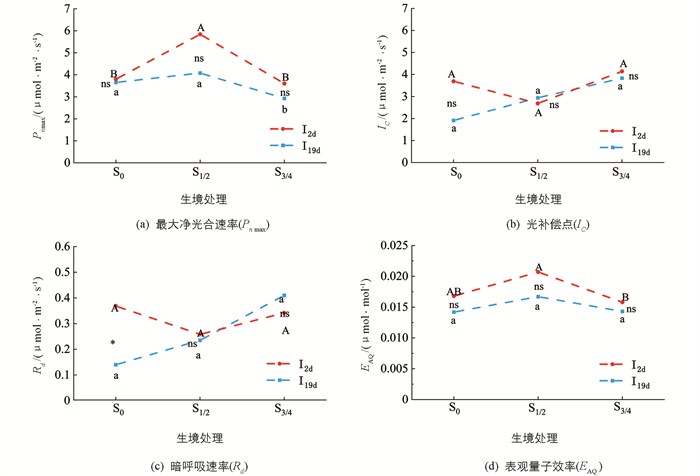
多重比较结果表明,在I2d降雨处理下,S1/2与S0和S3/4的Pnmax差异均具有统计学意义(p < 0.05),具体表现为S1/2 > S0 > S3/4. 在I19d降雨处理下,S3/4与S0和S1/2差异具有统计学意义(p < 0.05),具体表现为S1/2 > S0 > S3/4(图 3a).
t检验结果表明,无论是在哪种小生境中,I2d降雨处理与I19d降雨处理下桢楠幼苗的Pnmax差异均无统计学意义(图 3a).
双因素方差分析结果表明,降雨时间格局和垂直异质生境对桢楠幼苗Pnmax无交互作用(p > 0.05)(表 2).
-
单因素方差分析结果表明,无论是在I2d降雨还是在I19d降雨处理下,不同小生境之间桢楠幼苗的IC和Rd差异均无统计学意义(图 3b,3c).
t检验结果表明,无论是在哪种小生境中,I2d降雨与I19d降雨处理下桢楠幼苗的IC差异无统计学意义. 在S0小生境中,I2d降雨处理与I19d降雨处理下桢楠幼苗的Rd差异具有统计学意义,具体表现为I2d > I19d,在S1/2与S3/4小生境中差异无统计学意义(图 3b,3c).
双因素方差分析结果表明,降雨时间格局和垂直异质生境对桢楠幼苗IC及Rd无交互作用(表 2).
-
单因素方差分析结果表明,在I2d降雨处理下,S1/2小生境与S3/4小生境中桢楠幼苗的EAQ差异有统计学意义(p < 0.05);S0与S1/2之间,S0与S3/4之间差异无统计学意义,具体表现为S1/2 > S0 > S3/4. 在I19d降雨处理下,不同小生境之间桢楠幼苗EAQ差异无统计学意义(图 3d).
t检验结果表明,无论是在哪种小生境中,I2d降雨处理与I19d降雨处理下桢楠幼苗EAQ差异均无统计学意义(图 3d).
双因素方差分析结果表明,降雨时间格局和垂直异质生境对桢楠幼苗EAQ无交互作用(表 2).
2.1. 不同降雨时间格局下垂直异质生境中桢楠幼苗光响应过程
2.2. 光响应模型拟合优度评估
2.3. 光响应曲线特征参数比较
2.3.1. 最大净光合速率(Pnmax)
2.3.2. 光补偿点(IC)及暗呼吸速率(Rd)
2.3.3. 表观量子效率(EAQ)
-
光合作用是植物生长发育的基础,光响应曲线揭示了净光合速率与光合有效辐射之间的对应关系,由其反映出的光响应参数是评价光合作用的重要指标. 数学模型被广泛应用于光响应曲线的拟合中,不同模型的推导机理不同,拟合结果也不同. 从精度上来看,NRHM的R2最大,ERMS和EMA相对较小,拟合精度最高;MRHM,EM次之;RHM的精度最低,R2最小,ERMS和EMA较大,不适用于不同降雨时间格局下垂直异质生境中桢楠幼苗光响应曲线的拟合. 从拟合效果上看,RHM,NRHM,EM在定义的范围内均是严格单调的,没有最大值,无法准确描述出高光合有效辐射下曲线实际的降低走势. 而MRHM通过引入光抑制项β和光饱和项γ解决了其他3种模型无法拟合光抑制过程的缺陷[31-33],拟合精度高,由拟合曲线计算出的光响应参数值均非常合理. 由此可见,直角双曲线修正模型(MRHM)是本研究中桢楠光响应曲线的最适拟合模型,在部分C3和C4植物中也发现这一模型的拟合优度最佳[34].
-
植物扎根于土壤小生境,从中吸收水分、养分供叶片进行光合作用,所以不同土壤小生境大多在水分、矿质元素两因素的差异性上影响其植株的光合作用,可概括为两个方面:一是因土壤板结、岩溶裂隙增大入渗等原因导致土壤小生境中水分亏缺. 此状况下运输到叶片中的水分便也不充足,引起气孔关闭,进入叶片的CO2减少,而水和CO2均为光合作用的原料. 叶片缺水时,淀粉水解增强[35],糖类不断积聚,光合产物的输出变得缓慢,抑制了光合作用的速率;同时叶片缺水,叶绿体中电子传递速率降低,与光合磷酸化解耦联,影响同化力的形成[36];除此之外,缺水也会影响叶片的生长,光合面积扩展便也受到抑制,叶片吸收光能减少,这些都会导致光合速率的降低. 二是因土壤稀薄等原因导致土壤小生境中矿质营养亏缺. 例如,土壤中氧气含量不充足时,会阻碍根部的有氧呼吸,而根部是吸收矿质元素的机制之一,即主动吸收是借助根部细胞代谢提供的能量来进行的[35]. 根的有氧呼吸被抑制,吸收矿质元素的功能便也被抑制,矿质元素的缺乏,会在诸多方面对植物的光合作用产生影响. 比如N,Mg是叶绿素的组分,Mn,Zn,Cu等元素是其合成过程中某些酶的活化剂[35]. 缺乏这些矿质元素时,叶绿素合成受阻,植物对光能的收集、传递、转换便受到抑制,光合能力降低. 若严重缺氧,根通过无氧呼吸产生的酒精等毒害物质还会导致植物烂根,阻碍其发挥吸收、运输水分与养分的功能. 所以,妨碍了根系的活动,会间接对光合作用产生负面影响. 上述结果均会导致植物光合速率的降低.
光补偿点、暗呼吸速率和表观量子效率均能反映植物对弱光的利用能力. 结果表明,除I2d降雨下,S3/4小生境中桢楠幼苗的表观量子效率显著低于S1/2小生境外,垂直异质小生境对桢楠幼苗利用弱光的能力没有显著影响. 但所有处理中,桢楠幼苗的表观量子效率均较小,低于一般植物0.03≤EAQ≤0.05的水平[37],说明桢楠幼苗在弱光下的光能利用效率相对较低.
-
实验结果表明,延长降雨时间间隔并没有显著影响桢楠幼苗的光合潜能和利用弱光的能力,这可能是由于2种降雨格局下降雨总量相同而致. 仅在S0小生境中,不同降雨时间间隔下桢楠幼苗的暗呼吸速率差异有统计学意义,具体表现为I2d > I19d,说明随降雨时间间隔的延长,全土生境中的桢楠幼苗利用弱光的能力增强,但随着岩溶裂隙层的增厚,其漏失水养的负面影响使这种差异在其他2种小生境中不复存在.
3.1. 光响应曲线拟合模型的比较与评价
3.2. 垂直异质生境对桢楠幼苗光响应参数的影响
3.3. 不同降雨时间格局对桢楠幼苗光响应参数的影响
-
不同数学模型对不同降雨时间格局下喀斯特垂直异质生境中桢楠幼苗净光合速率的光响应过程及参数的拟合存在一定差异,MRHM更适用于拟合该生境中桢楠幼苗的光响应曲线,不但与实测值相差很小,而且可以准确地拟合出达到饱和光合有效辐射之后的光抑制现象,具有较高的拟合优度. 垂直异质生境显著影响桢楠幼苗的最大净光合速率,反映出岩溶裂隙层具有一定的生境功能. 各生境中桢楠幼苗在不同降雨时间间隔下的光合潜力及弱光利用能力差异无统计学意义,这表明桢楠能较好地适应生境水分亏缺,可作为西南喀斯特地区石漠化山地植被恢复重点考虑的树种.