


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
大豆(Glycine max)起源于中国,是双子叶植物纲蔷薇目豆科豆属的一年生草本植物,至今已有5 000年左右的栽培耕作和食用历史. 大豆目前作为全球重要的粮油作物之一,供给全球70.86%的植物蛋白和28.88%的植物油脂[1]. 2020年,我国大豆播种面积仅次于玉米、稻谷和小麦. 国家统计局数据显示,2020年我国进口大豆首次突破1亿t,占当年国内大豆总消耗量的90%.
大豆主要用途是榨油,不饱和脂肪酸中的亚油酸和亚麻酸具有功能性保健作用. 大豆粗脂肪中的不饱和脂肪酸包括棕榈油酸、油酸、亚油酸、亚麻酸和二十碳烯酸,其中亚油酸和油酸含量最高,分别占不饱和脂肪酸的35%~60%和20%~50%[2]. 我国大豆含油量较低且胞内存在占比较大的蛋白质,使得其油脂提取困难[3],提高我国大豆粗脂肪含量十分必要.
大豆品质性状粗脂肪含量属多基因控制的数量性状,易受环境影响. 现代分子标记的发展为改良数量性状提供了一种快速有效的方法. 高密度遗传连锁图谱的构建为大豆重要农艺性状的数量性状位点(Quantitative Trait Locus,QTL)挖掘奠定了基础,但已经公布的大豆蛋白质含量和脂肪含量相关QTL与其连锁标记距离较远,还不能满足分子标记辅助选择和图位克隆的要求. 本研究利用高品质大豆品种长江春2号[4]与适合重庆气候条件的鲜食大豆品种渝蜀鲜2号为亲本,建立了春大豆品种间杂交(长江春2号×渝蜀鲜2号)F2群体. 采用SSR和In-del标记,在构建该群体高密度遗传连锁图谱的基础上,结合F2单株和F2∶3,F2∶4,F2∶5共4个世代的粗脂肪含量,对大豆粗脂肪含量QTL进行定位,并筛选含有增加粗脂肪含量等位基因的株系,为进一步实现大豆粗脂肪含量QTL的精细定位和图位克隆奠定基础.
全文HTML
-
大豆品种长江春2号由重庆市农业科学院和自贡市农业科学研究所选育,该品种属于高蛋白中熟常规大豆,综合性状优良. 渝蜀鲜2号是经重庆市农业科学院和四川省农业科学院经济作物研究所合作审定的鲜食大豆.
本研究的材料为长江春2号(母本)与渝蜀鲜2号(父本)杂交得到的F2代分离群体,2021年在重庆播种后就已经采用Zhang等[5]改良后的CTAB法提取了该群体186株单株DNA用于后续试验. 此外,在2022年重庆、2022年云南和2023年重庆分别播种后收获了F2∶3,F2∶4,F2∶5代的种子,并测出两个亲本及其4个不同环境下大豆种子的粗脂肪含量用于后续定位.
-
本研究使用FOSS NIRS DS 2500对两个亲本以及F2,F2∶3,F2∶4,F2∶5等材料进行粗脂肪含量测定.
-
通过SPSS软件对亲本和上述4个不同环境下的大豆粗脂肪含量进行统计分析,计算出平均值、最大值、最小值、偏度、峰度和变异系数等统计指标. 此外,利用该软件对这些性状进行相关性分析,以探究它们之间的关系. 同时,使用Origin 2019软件来绘制这些性状的频率分布直方图,以便更直观地展示数据分布情况.
-
本项研究中所使用的简单重复序列(Simple Sequence Repeats,SSR)引物序列是从大豆标记的公共数据库Soybase(
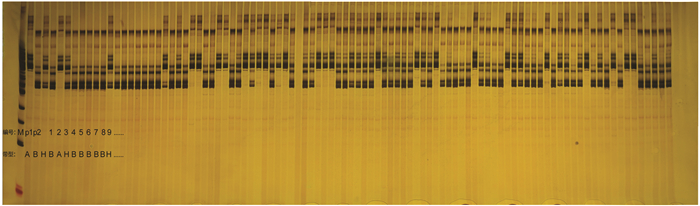
http://www.soybase.org/ )中下载获取的. 同时针对两个亲本品种设计插入和缺失(Insertion and Deletion,InDel)引物,SSR和InDel引物由上海生工生物工程有限公司合成. 从大豆的20条染色体中挑选出3 780对均匀分布的SSR引物,用于筛选具有多态性的引物对. 这些多态性引物被用于对(长江春2号×渝蜀鲜2号)F2群体进行标记基因型的检测. 在数据统计过程中,将带型与长江春2号相同的标记为A,与渝蜀鲜2号相同的标记为B,出现杂合型(同时具有A和B带型)的标记为H,缺失带型的标记为U. -
利用JoinMap 4.0软件作图,采用LOD值为4.0和重组率阈值为0.4的标准,同时应用了Kosambi作图函数进行遗传连锁分析[6].
-
在本项研究中,利用MapQTL 6.0软件对大豆的品质性状进行了QTL定位以及效应的检测. 当LOD值超过3.0时,判定为存在粗脂肪含量QTL. 在评估加性效应时,以渝蜀鲜2号为遗传背景,其中正效应意味着长江春2号的等位基因会增加性状的表型值,而负效应则表示长江春2号的等位基因会减少性状的表型值.
QTL命名方式定为“QTL+性状+染色体+数字”,其中以小写字母q开头表示QTL,英文缩写表示性状,后续数字表示该QTL位于哪条染色体,最后的数字则表示同一性状在该连锁群上检测到的不同QTL的个数. 例如:qOIL14.2表示在14号染色体上与粗脂肪含量相关的第2个QTL.
1.1. 试验材料
1.2. 粗脂肪含量的测定
1.2.1. 测定方法
1.2.2. 表型数据统计分析
1.3. 遗传连锁图谱构建
1.3.1. 多态性引物筛选与群体标记基因型检测
1.3.2. 遗传连锁图谱构建
1.4. 粗脂肪(OIL)含量QTL定位
-
在2021年到2023年的4个环境中,分别对两个亲本以及F2,F2∶3,F2∶4,F2∶5群体进行粗脂肪含量表型性状鉴定(表 1).
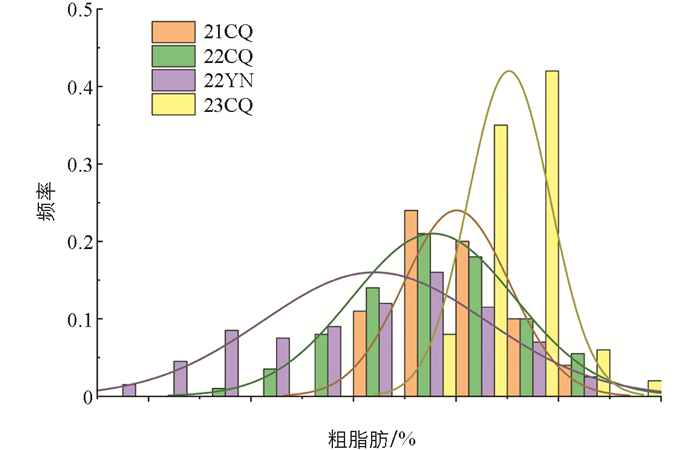
数据显示,2021年重庆,群体粗脂肪含量的平均值是20.01 g,在18.07~22.21 g范围内;2022重庆,群体粗脂肪含量的平均值是19.22 g,在15.15~22.92 g范围内;2022年云南,群体粗脂肪含量的平均值是17.24 g,在13.58~22.73 g范围内;2023年重庆,群体粗脂肪含量的平均值是21.04 g,在19.20~23.61 g范围内. 2021年重庆和2023年重庆两个亲本的粗脂肪含量相近,而2022年重庆和云南出现了两个亲本表型差异较大的现象. 2021年重庆,粗脂肪含量平均值略低于两个亲本,但差距很小,其余3个环境下群体的粗脂肪含量都位于两个亲本之间. 大豆脂肪含量是复杂的数量性状,受主效和微效的多基因控制[7],在已经报道的大豆粗脂肪含量QTL中,绝大多数都不能在多个环境中被检测到[8-10],本研究在不同环境间粗脂肪含量的平均值和变幅都存在一定差异,这也是造成粗脂肪含量QTL定位结果不稳定的原因.
对各个性状偏度和峰度的分析揭示了粗脂肪含量的分布接近正态分布(表 1). 性状的表型值频率分布直方图也显示了性状的连续性和近似正态分布(图 1),这与由多个基因控制的数量性状的遗传特性相符. 此外,性状的频率分布情况随环境的不同而变化,这表明它们都受到环境因素的影响.
-
本研究共计利用3 780对SSR引物对亲本长江春2号和渝蜀鲜2号进行引物多态性筛选,获得493对多态性引物,多态性比例为13.04%. 不同引物间的多态性差异很大. 其中Sat引物多态性比例最高,占比24.92%,其次为CSSR引物,占比24.14%,后面依次为Satt引物,占比16.28%;SSR引物,占比15.56%;GMES引物,占比14.40%;SWU引物多态性比例最低,仅占比10.18%,具体数据如表 2所示.
-
利用493对(表 2)经过筛选的对两亲本具有多态性的SSR引物,以及针对亲本品种长江春2号和渝蜀鲜2号设计的86对Indel引物,对F2群体中的186株单株进行了标记基因型的检测(图 2),共发现了518个具有多态性的位点.
-
在实验室前期构建的遗传图谱基础上[9],通过JoinMap 4.0分析软件,对518对具有多态性的标记位点进行了遗传连锁分析,并构建了长江春2号和渝蜀鲜2号之间的遗传连锁图谱(图 3). 该图谱的总长度为2 469.3 cM,包含了20个连锁群,这些连锁群均匀分布在大豆20条染色体上,每条染色体上标记数目差异较大. 图谱中共包含480个标记(表 3),标记间的平均距离为5.14 cM. 在图谱中,覆盖度最大的是第13号染色体(Chr.13),其总长度为200.30 cM,包含了44个标记位点,是标记数目最多的连锁群,且其标记间的平均距离为4.55 cM,覆盖度最小的为5号染色体(Chr.5),总长度为29.80 cM,包含7个标记位点,标记间的平均距离为4.26 cM.
-
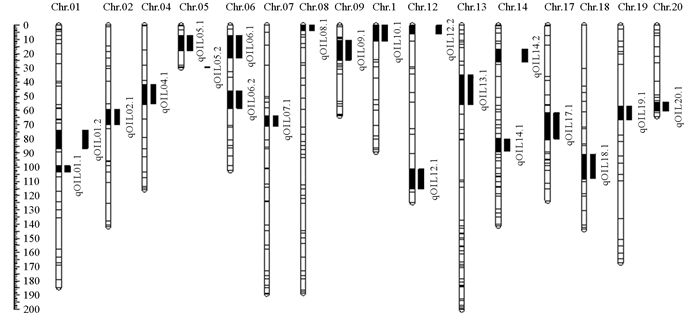
定位结果如表 4所示,共检测到21个粗脂肪含量(OIL)QTL,除了3,11,15,16号染色体(Chr.3,Chr.11,Chr.15,Chr.16)以外,其余16条大豆染色体上均检测到了不少于一个与粗脂肪含量相关的QTL,LOD值大小为3.00~8.08,表型贡献率为7.20%~18.10%. 定位到的粗脂肪含量相关的QTLs在染色体上的位置如图 4所示
在这些QTL中,LOD值最高的是位于2023年重庆环境的qOIL01.2,其LOD值为8.08,解释粗脂肪含量表型变异率为18.10%,长江春2号的等位基因的加性效应表现为增加大豆粗脂肪含量0.47%. 在这些QTL中,有3个(qOIL01.1,qOIL04.1,qOIL14.2)在两个不同的环境中被检测到. qOIL01.1在2022年重庆和2022年云南的LOD值分别为3.00和3.72,分别解释了7.20%和9.10%的表型变异,其中2022年重庆长江春2号等位基因的加性效应表现为增加表型值0.57%,而2022年云南表现为降低表型值0.44%. qOIL04.1在2021年重庆和2022年云南的LOD值分别为3.42和3.29,分别解释了10.80%和8.10%的表型变异,长江春2号的等位基因加性效应均表现为减少表型值. qOIL14.2在2022年重庆和2023年重庆的LOD值分别为5.78和4.18,分别解释了13.30%和9.80%的表型变异,2022年重庆长江春2号等位基因的加性效应表现为增加表型值0.48%,而2023年重庆表现为降低表型值0.28%.
2.1. 亲本及群体粗脂肪含量表型分析
2.2. 引物多态性筛选
2.3. 群体基因型检测
2.4. 遗传图谱的构建
2.5. 粗脂肪含量QTL定位
-
大豆遗传图谱的早期研究起步晚、发展速度慢,随着分子标记技术的发展,遗传学家开始运用SSR等分子标记技术辅助构建大豆遗传图谱[12],Akkaya等[13]证实了SSR标记在大豆基因组中的多态性. 本研究除了利用SSR引物,还设计出一套针对(长江春2号×渝蜀鲜2号)F2群体的InDel引物,使得构建的图谱具有更丰富、更准确的标记.
本课题组前期已经开展大豆种子大小以及品质性状的QTLs定位研究,最终筛选出了18个可能参与种子大小调控的候选基因[14]和32个品质相关的候选基因[15]. 自Cregan等[16]构建了第一张大豆的公共遗传连锁图谱以来,利用高密度遗传连锁图谱进行QTL定位的研究陆续开展[17-18]. 本研究基于对构建的遗传图谱进行加密,结合F2和F2∶3,F2∶4,F2∶5 4个世代粗脂肪含量表型数据进行粗脂肪含量的QTL定位. 目前我们成功构建的大豆遗传连锁图谱具有更高的标记密度和更大的覆盖度,相邻标记之间的距离相对较短,有助于提高QTL定位的精度.
通过检测在不同遗传背景和环境条件下稳定表达的QTL,利用与目标性状基因连锁的分子标记来筛选目标性状基因型,可以提高QTL的跟踪效率,缩短育种周期,提高育种效率[19]. 不同环境下均检测出来的QTL为稳定QTL. 本研究结果显示,其中3个QTLs(qOIL01.1,qOIL04.1和qOIL14.2)在两个环境中均能检测到,增强了这些QTLs与粗脂肪含量的关联可靠性,更有可能为未来的育种工作提供重要的候选基因.
本研究所用的亲本材料长江春2号是重庆市审定的高蛋白大豆品种中脂肪含量最高的品种,渝蜀鲜2号是重庆市审定的鲜食大豆品种中脂肪含量较低的品种,两个亲本间的粗脂肪含量差异显著,是挖掘粗脂肪含量候选基因理想的亲本品种. 另外,长江春2号具有种子产量高和品质优等特点,渝蜀鲜2号具有种子百粒质量高和发芽率高等特点,两个品种配制的杂交分离后代,是较好开展产量、品质性状基因定位和分子标记辅助选择的群体材料.
本研究旨在更深入地探索大豆粗脂肪含量的遗传基础,为下一步挖掘出粗脂肪含量候选基因、基因克隆和功能验证奠定坚实基础,为我国大豆育种中的粗脂肪含量提升提供有效分子标记和遗传资源.