



 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
大豆(Glycine max)是豆科大豆属一年生草本植物,其籽粒是植物蛋白和油脂的重要来源. 我国是大豆原产国,具有丰富的种质资源,但大豆消费严重依赖进口. 提高种质资源利用效率,增加大豆产量,对于促进大豆相关产业可持续发展有重要意义. 在“绿色革命”中,利用水稻和小麦矮生资源选育半矮化品种,提高种植密度,使作物产量急剧增加[1]. 其中,水稻矮生资源sd1是赤霉素(Gibberellins,GAs)合成酶基因GA20ox的缺失突变体[2];小麦矮生资源rht1是DELLA基因功能获得性突变体[1],这两种突变体均促进了DELLA蛋白的累积,使株高降低. 因此,DELLA基因家族有重要的应用价值.
DELLA蛋白是GAs信号传导途径中的一类负调控因子,其N端具有DELLA结构域,包含DELLA,LExLE,TVHYNP 3段保守序列,是GAs受体蛋白GID1(Gibberelin Insensitive Dwarf1)的识别位点[3-4];C端具有GRAS蛋白家族的特征,包含LR1,NLS,VHIID,LR2,SH2-like,PFYRE,RVER和SAW等保守区,主要与顺式调控元件结合和其他调控因子互作. 在GAs的信号传导途径中,当GAs与GID1结合后,GID1的构象改变,进入细胞核作用于DELLA蛋白[3, 5-6]. GAs-GID1-DELLA复合物与SCF E3泛素连接酶作用,使DELLA蛋白通过泛素化途径降解,启动下游基因表达[7]. 当GAs浓度很低时,DELLA蛋白在细胞中累积,植物表现出矮化等性状[8].
在拟南芥中发现了5个DELLA基因,即GAI,RGA和RGL1/2/3[8-11],其中GAI和RGA的获得性突变体极度矮化;水稻DELLA基因缺失突变体slr1(slender1)表现出细长的表型[12];玉米d8,d9突变体极度矮化[1];小麦RHT1基因获得一系列突变体(如rht-B1d,rht-B1b,rht-D1b,rht-D1d,rht-B1e和rht-B1c等)表现出高矮不一的表型[13]. 除了对株高的调控外,DELLA基因还参与植物多方面的生长发育调控. 葡萄L1基因编码的DELLA序列突变为DELHA,使葡萄蔓藤上的蔓须变成了花序[14];在拟南芥中的研究也认为RGA,RGL1和RGL2基因参与花器官发育调控[15],转录因子WRKY75和GAI,RGA蛋白互作调控植株开花[16];RGL1和RGL2参与调控种子萌发[17]. 这些研究报道表明DELLA基因与株高等多种重要性状相关.
最近,在大豆中报道了4个DELLA基因,即GmRGAa/b和GmRGLa/b. 其中,GmRGAs能够正调控转录因子GmSTF1/2的蛋白水平,而GmSTF1/2通过激活赤霉素代谢基因GmGA2ox-7b的表达使大豆株高降低[18]. 为了进一步了解大豆DELLA基因的遗传变异,本文利用29个大豆种质资源的基因组数据,用生物信息学的方法鉴定了232个大豆DELLA基因家族成员,并分析了其生物学功能.
全文HTML
-
在大豆SoyOmics[19]数据库下载28个大豆种质资源的基因组数据,资源名称分别是Xu Dou No.1,Tie Feng No.18,Ju Xuan No.23,Wan Dou No.28,Amsoy,Yu Dou No.22,Jin Dou No.23,Qi Huang No.34,Han Dou No.5,PI 548362,Ji Dou No.17,Dong Nong No.50,Hei He No.43,Ke Shan No.1,Zhutwinning2,Zi Hua No.4,Tong Shan Tian E Dan,58-161,PI 398296,Zhang Chun Man Cang Jin,Feng Di Huang,Tie Jia Si Li Huang,Shi Sheng Chang Ye,PI 562565,PI 549046,PI 578357,W05,Zhonghuang 13,对应编号为SoyC01,SoyC02,SoyC03,SoyC04,SoyC05,SoyC06,SoyC07,SoyC08,SoyC09,SoyC10,SoyC11,SoyC12,SoyC13,SoyC14,SoyL01,SoyL02,SoyL03,SoyL04,SoyL05,SoyL06,SoyL07,SoyL08,SoyL09,SoyW01,SoyW02,SoyW03,W05,ZH13. 其中,SoyC表示栽培种,SoyL表示地方种,SoyW01,SoyW02,SoyW03,W05为野生大豆(Glycine soja Sieb. and Zucc.). 在Phytozome数据库中下载大豆Williams82 (Wm82)的基因组数据.
-
拟南芥DELLA蛋白GAI,RGA,RGL1,RGL2和RGL3的蛋白序列从拟南芥TAIR数据库中下载. 分别用这5个DELLA蛋白序列在Phytozome数据库中检索Wm82的基因组,筛选比对长度接近蛋白全长,编码蛋白序列一致性在45%以上的基因. 候选的大豆DELLA基因编码的蛋白序列在SMART蛋白数据库中进一步分析,鉴定是否属于DELLA蛋白家族成员.
-
在大豆SoyOmics数据库中,用Wm82的DELLA基因分别检索28个种质资源的基因组数据,筛选覆盖度在85%以上,E值为0的基因,作为各大豆种质资源的DELLA基因. 用MegAlign软件分别对所有DELLA基因的DNA和蛋白质序列进行多序列比对和进化树分析.
-
用MegAlign软件对Wm82的DELLA蛋白序列进行比对,参照文献报道[20]对DELLA蛋白的保守结构域进行比较.
-
从SoyOmics数据库中查询Zhonghuang 13(ZH13)的转录组数据,根据FPKM值在图云平台(
http://cloudtutu.com.cn/ )绘制基因表达热图. -
用Wm82的DELLA蛋白序列在STRING[21]数据库中搜索存在互作关系的大豆蛋白,最低交互得分设定为0.400.
1.1. 大豆种质资源基因组数据
1.2. 大豆DELLA基因的鉴定
1.3. 大豆DELLA基因的泛基因组分析
1.4. 大豆DELLA蛋白质结构分析
1.5. 大豆DELLA基因表达模式分析
1.6. 蛋白互作网络分析
-
以拟南芥的5个DELLA蛋白序列在Phytozome数据库中搜索,从大豆Wm82的基因组中找到8个DELLA基因. 这8个DELLA基因分别位于11,18,5,8,10,20,4和6号染色体,基因ID号为Glyma.11G216500,Glyma.18G040000,Glyma.05G140400,Glyma.08G095800,Glyma.10G190200,Glyma.20G200500,Glyma.04G150500和Glyma.06G213100,命名为GmGAI1 (Wm82.Chr11),GmGAI1 (Wm82.Chr18),GmGAI2 (Wm82.Chr05),GmGAI2 (Wm82.Chr08),GmRGL1 (Wm82.Chr10),GmRGL1 (Wm82.Chr20),GmRGL2 (Wm82.Chr04)和GmRGL2 (Wm82.Chr06). 序列分析表明,GmGAI1 (Wm82.Chr11),GmGAI1 (Wm82.Chr18),GmGAI2 (Wm82.Chr05)和GmGAI2 (Wm82.Chr08) 与拟南芥GAI和RGA的同源性较高,编码蛋白质序列的一致性为55.8%~64.9%. GmRGL1 (Wm82.Chr10),GmRGL1 (Wm82.Chr20),GmRGL2 (Wm82.Chr04)和GmRGL2 (Wm82.Chr06)的编码蛋白与拟南芥RGL1/2/3蛋白的一致性在37.1%~43.8%. 用Wm82的8个DELLA基因在数据库SoyOmics中查询,从4个野生大豆、9个地方品种和15个栽培种中,分别找到32,72和120个大豆DELLA基因.
对232个大豆DELLA基因进行系统发生树分析(图 1),发现GmGAI1基因可分为两个亚组,来源于18号染色体的GmGAI1组成一个亚组,在这一亚组中野生大豆SoyW01,SoyW02,SoyW03和W05的GmGAI1在进化树末端;来源于27个种质资源11号染色体的GmGAI1基因和来源于2个野生大豆SoyW02,W05的13号染色体的GmGAI1基因组成另一个亚组,这2个野生大豆的GmGAI1基因和其他种质资源GmGAI1基因的遗传距离相对较远. 同样的,来源于5号和8号染色体的GmGAI2基因分别组成2个亚组,4个野生大豆的GmGAI2基因处于靠近进化树末端的位置. 来源于4号、6号、10号和20号染色体的GmRGL1和GmRGL2基因分别组成4个亚组,每个亚组再分2~3个小亚组,整体上野生大豆的GmRGL1和GmRGL2基因与地方种和栽培种的GmRGL1和GmRGL2基因遗传距离相对较远. 这一结果初步表明大豆野生种驯化为地方种和栽培种的过程中,DELLA基因受到了选择;地方种和栽培种11号染色体上的GmGAI1基因可能由野生大豆SoyW02和W05的13号染色体上的GmGAI1基因发生交换而来.
-
比较232个大豆DELLA基因的开放阅读框(Open Reading Frames,ORFs),发现都只有一个外显子,其中有19个DELLA基因发生了移码突变,其中13个基因不能编码有DELLA蛋白特征的产物,分别为GmGAI1 (SoyL06.Chr11),GmGAI2 (SoyC08.Chr08),GmGAI2 (SoyC01.Chr08),GmGAI2 (SoyL02.Chr08),GmRGL1 (SoyL05.Chr20),GmRGL1 (SoyC06.Chr20),GmRGL1 (SoyW03.Chr20),GmRGL1 (SoyL01.Chr20),GmRGL1 (SoyL02.Chr20),GmRGL1 (SoyC08.Chr20),GmRGL1 (SoyC08.Chr04),GmRGL2 (SoyC13.Chr06)和GmRGL2 (SoyC06.Chr06);有6个基因编码的DELLA蛋白在C端提前终止,分别为GmGAI1 (SoyL01.Chr11),GmGAI2 (SoyC02.Chr05),GmGAI2 (SoyC12.Chr08),GmRGL1 (SoyL07.Chr10),GmRGL2 (SoyC14.Chr06)和GmRGL2 (SoyL06.Chr06).
根据ORFs预测了219个DELLA基因的蛋白序列,通过同源性分析大豆DELLA蛋白可分成8个亚组,整体上源于相同染色体的DELLA蛋白属于一个亚组,其中GmGAI1s和GmGAI2s的同源性更近,GmRGL1s和GmRGL2s的同源性更近. 在GmGAI1s蛋白亚组中,源于野生大豆SoyW02和W05的13号染色体和其他种质资源11号染色体的GmGAI1蛋白在一个亚组. 除了在C端提前终止的DELLA蛋白外,每个亚组中DELLA蛋白的序列一致性在98.8%~100%. 4个野生大豆中源于4号染色体的GmRGL2蛋白与亚组中其他种质的GmRGL2蛋白序列高度一致,而野生种源于其他染色体的DELLA蛋白与同组蛋白的序列一致性相对更远.
由于每个亚组的大豆DELLA蛋白序列高度一致,选取Wm82的8个DELLA蛋白进行分析,比较不同亚组间DELLA蛋白的序列差异(图 2). 比对结果显示,GmGAI1(Wm82.Chr11)和GmGAI1 (Wm82.Chr18),GmGAI2 (Wm82.Chr05)和GmGAI2 (Wm82.Chr08),GmRGL1 (Wm82.Chr10)和GmRGL1 (Wm82.Chr20),GmRGL2 (Wm82.Chr04)和GmRGL2 (Wm82.Chr06)间的序列一致性分别为95.4%,93.6%,91.0%和94.6%. 蛋白结构分析表明,GmGAI1 (Wm82.Chr11),GmGAI1 (Wm82.Chr18),GmGAI2 (Wm82.Chr05)和GmGAI2 (Wm82.Chr08)的DELLA结构域有完整的DELLA,LExLE和TVHYNP保守序列;而GmRGL1 (Wm82.Chr10),GmRGL2 (Wm82.Chr04)和GmRGL2 (Wm82.Chr06)的DELLA,LExLE和TVHYNP保守序列与GmGAI1s和GmGAI2s的不一致. 值得注意的是,GmRGL1 (Wm82.Chr20)的DELLA结构域发生缺失. 这些结果暗示大豆的8个DELLA基因在生物学功能上有差异,与拟南芥GAI蛋白同源性高的4个GmGAI1s和GmGAI2s蛋白在功能上可能冗余.
-
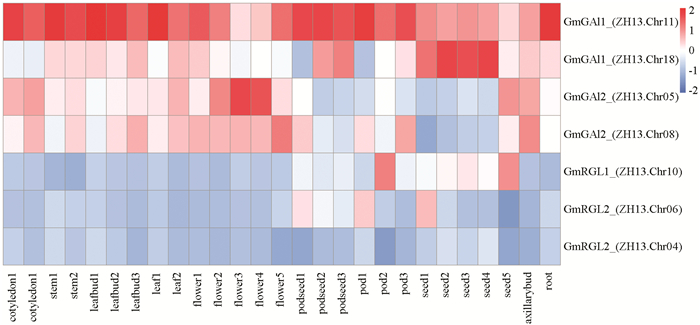
在大豆数据库SoyOmics中,通过转录组测序,获得了ZH13不同发育时期植物组织的基因表达数据. 以此为基础,绘制大豆DELLA基因在根、茎、叶、花、果等不同组织部位的表达模式热图(图 3),根据基因表达强弱预测DELLA基因在大豆生长发育不同阶段的生物学功能. 从分析情况来看,GmGAI1 (ZH13.Chr11)在大豆不同发育阶段的组织器官中均高表达;GmGAI1 (ZH13.Chr18)在大豆不同发育阶段的籽粒中优势表达;GmGAI2 (ZH13.Chr05)在花中优势表达;GmGAI2 (ZH13.Chr08)在成熟的茎、叶、花、腋芽等部位表达,但在早期发育的籽粒表达水平较低;GmRGL1 (ZH13.Chr10)和GmRGL2 (ZH13.Chr06)分别在不同发育时期的籽粒和果荚中表达,在其他植物组织中表达水平较低;GmRGL2 (ZH13.Chr04)在所有植物组织中表达水平均较低. 而GmRGL1 (ZH13.Chr20)没有基因表达数据. 这些结果表明大豆DELLA基因表达模式存在差异,可能在植物组织器官发育过程中发挥不同的功能.
为了解上游调控序列差异对基因表达情况的影响,分别对GmGAI1 (ZH13.Chr11)和GmGAI1 (ZH13.Chr11Chr18),GmGAI2 (ZH13.Chr05) 和GmGAI2 (ZH13.Chr08),GmRGL2 (ZH13.Chr06)和GmRGL2 (ZH13.Chr04)这3对基因的起始密码子上游1500bp的序列进行了分析和比较,两两间基因上游调控序列的一致性分别为79.0%,77.2%和70.1%. 在New PLACE数据库分析,显示GmGAI1 (ZH13.Chr11),GmGAI1 (ZH13.Chr11Chr18),GmGAI2 (ZH13.Chr05),GmGAI2 (ZH13.Chr08),GmRGL2 (ZH13.Chr06)和GmRGL2 (ZH13.Chr04)的上游调控序列分别有83,77,83,84,79和81种不同的顺式调控元件,其中6者共同有的顺式调控元件有37种. 同一亚组的DELLA基因上游调控序列中顺式元件种类的差异如表 1所示. 从表中所示的差异调控元件来看,同一亚组的两个DELLA基因在转录水平可能受到不同的调控,如GmGAI1 (ZH13.Chr11)的上游调控序列中MYB2CONSENSUSAT元件与干旱应答和ABA信号有关,MYBPLANT元件与花发育调控有关;GmGAI1 (ZH13.Chr18)的上游调控序列中AMYBOX1元件与淀粉酶基因表达有关. 这些数据暗示大豆DELLA基因在适应环境和组织器官发育调控过程中发挥了不同的功能.
-
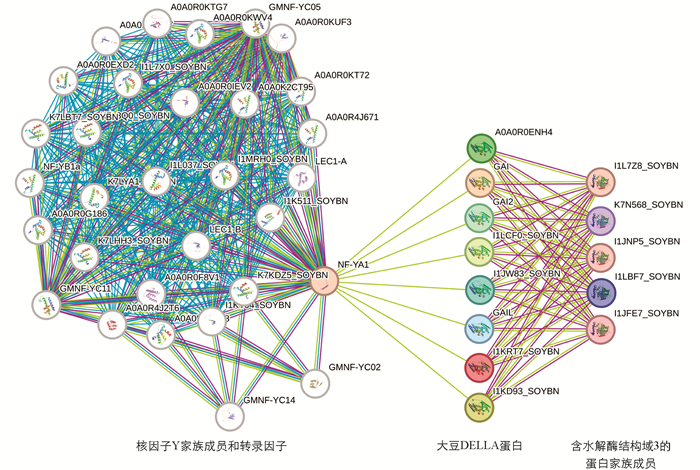
在STRING数据库中,用GmGAI1 (Wm82.Chr11),GmGAI1 (Wm82.Chr18),GmGAI2 (Wm82.Chr05),GmGAI2 (Wm82.Chr08),GmRGL1 (Wm82.Chr10),GmRGL1 (Wm82.Chr20),GmRGL2 (Wm82.Chr04)和GmRGL2 (Wm82.Chr06)的蛋白序列搜索在大豆中能发生互作的蛋白. 如图 4所示,大豆DELLA蛋白能与5个含水解酶结构域3的蛋白(Abhydrolase_3 domain-containing protein,ABHD3)互作,也能与核因子NF-YA1(nuclear factor Y-A1)蛋白互作. 而核因子NF-YA1能够与其他核因子家族成员NF-YB、NF-YC以及转录因子LEC1-A/B等互作,激活或者抑制下游基因表达. 这些互作蛋白涉及到多种生物学过程,包括赤霉素信号传导、花器官形态建成、氧自由基信号响应、干旱应答、脂质代谢应答、棉子糖家族低聚糖生物合成等. 分析结果与报道的DELLA蛋白是赤霉素信号传导负调控因子相符合,并进一步预测是通过与核因子NF-YA1互作来调控下游基因表达.
2.1. 大豆种质资源DELLA基因家族成员的鉴定和系统发生树分析
2.2. 大豆种质资源的DELLA基因结构和蛋白序列分析
2.3. 大豆DELLA基因在不同组织部位的表达和上游顺式调控元件分析
2.4. 大豆DELLA蛋白互作网络分析
-
我国大豆单产偏低,增加种植密度提升单产,是当前提高大豆产量的一个重要途径. DELLA基因是赤霉素信号的负调控因子,参与了调控植物的株高和避荫性反应. 在绿色革命中,小麦RHT1基因不同类型的突变体出现一系列高矮不一的表型[1, 13],利用这些种质资源选育耐密植品种,增加种植密度,大幅提升了小麦产量. 因此,对大豆DELLA基因开展深入研究,创制耐密植的大豆种质资源,具有重要的意义.
本研究利用29个大豆种质资源的基因组数据,分析比较大豆种质资源间DELLA基因的亲缘关系和遗传变异情况,发现4个野生大豆在4号染色体上的GmRGLs基因编码的蛋白序列一致性较高,暗示这个基因在功能上可能比较保守. 而野生大豆中其他的DELLA基因和地方种、栽培种的相比遗传距离相对较远,表明这些基因在驯化过程中受到了选择. DELLA基因的这些遗传变异是否与野生大豆从藤状生长形态转变为直立生长形态有关,还需要深入研究. 同时,部分种质资源的DELLA基因发生了无义突变和翻译提前终止的移码突变,这些突变是否与种质资源株高、结实性等性状的变异相关,也需要进一步探索.
赤霉素调控了植物株高、开花、结实、根伸长等生长发育过程,而DELLA蛋白是赤霉素信号传递途径中的核心因子,大豆DELLA基因的表达模式与赤霉素调控生长发育的作用相符合. 对大豆DELLA基因的表达情况分析表明,位于11号染色体上的GmGAI1基因在所有植物组织中均高表达,可能发挥了主要作用,而11号染色体上的GmGAI1基因表达量稍弱,可能发挥次要作用,这与最近报道的GmRGAb对株高的调控作用要强于GmRGAa的结论一致[18]. GmRGL1 (ZH13.Chr10)和GmRGL2 (ZH13.Chr06) 分别在发育中的籽粒和果荚中优势表达,这两个基因在籽粒和果荚发育中的功能还有待深入鉴定. 同时,通过蛋白互作分析,发现大豆DELLA蛋白能够与ABHD3和NF-Y等蛋白家族互作,这里面部分蛋白已被证明与花器官发育、逆境应答等途径相关,筛选的这些互作蛋白有利于进一步揭示DELLA蛋白的作用机理.
综合而言,本研究利用29个大豆种质资源的基因组数据信息,对大豆DELLA基因进行泛基因组分析,初步明确DELLA基因的遗传变异和进化关系;利用转录组数据对大豆DELLA基因在不同植物组织器官的表达情况进行了分析比较,推测大豆DELLA基因在植物生长发育的各阶段发挥不同的生物学功能;通过蛋白互作分析,为揭示DELLA蛋白的作用机制提供了新的候选因子. 这为明确大豆DELLA蛋白的生物学功能以及创制耐密植大豆种质提供了参考.