


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
大豆是全球重要的粮食、经济作物,具有丰富的蛋白质和油分,全球超过56%的油脂、25%的人类食品及动物饲料所用蛋白由大豆提供[1]. 大豆原产于我国,早在5000年前,我国就已经开始栽培大豆[2],但随着我国经济的不断发展,城市建筑征用的土地面积逐渐增大,耕地面积大量减少,且大豆与玉米、水稻等作物相比,产量和效益都没有优势,农民种植大豆的积极性不高,大豆年产量严重不足. 然而我国大豆消费需求量大,平均每年所需总量约1.1亿t,其中进口9 000万t左右,对国外输入依赖程度较高,达80%以上[3]. 为了促进我国大豆产业的发展,保障粮食安全,应该扩大种植面积,增加大豆产量.
川西高原地区总面积约为23.6万km2,其中约有一半的面积可以开发用作耕地. 但当地气温低,冬季长,主要农作物青稞的生物量小,营养价值低,导致牦牛冬春季饲草短缺,掉膘严重,死亡率高[4]. 玉米、大豆饲用价值高,可以作为优质原料进行青贮饲料的生产,其中高原豆类种植鲜质量可超过80 t/hm2,产量前景乐观[5]. 因而,玉米、大豆等做成的青贮饲草能有效缓解川西高原地区饲草原料短缺问题. 且川西高原地区日照条件充足,适宜进行玉米、大豆带状间作种植,但当地气温低、昼夜温差大,大豆易遭受低温胁迫,导致大豆生育期延后,株高、叶片鲜质量、茎鲜质量显著降低[6],光合作用的性能和活性被抑制[7]. 此外,在川西高原地区进行玉米、大豆带状间作种植过程中,高位作物玉米会对低位作物大豆产生荫蔽胁迫,影响大豆植株的生长和光合特性[8],使其光合能力及光合产物合成受到抑制,植株茎秆纤细、陡长、易倒伏,产量显著下降[9].
综上所述,在已有研究的基础上,本文旨在探究低温和荫蔽双重胁迫对大豆光合特性的影响,为川西高原地区乃至全国进行玉米、大豆带状间作种植提供一定的依据.
全文HTML
-
大豆:中黄311(课题组前期研究表明该品种适宜高海拔地区进行净作种植),共处理120盆.
-
本试验在四川农业大学成都校区第三教学楼的恒温培养箱中进行,为双因素随机试验,因素一为温度(T),T1为正常温度、T2为低温;因素二为光环境(L),L1为正常光、L2为荫蔽. 各处理的温度、光强等数据见表 1. 选择若干外形完整、大小均一且饱满的大豆种子,用75%的酒精浸泡2~3 min消毒,在流水下冲洗至无酒精味后,将其放在打湿的纱布上进行暗萌发. 待长出1 cm左右胚根后,移栽至7 cm×7 cm×10 cm(长×宽×高)的花盆中. 每隔1 d施用0.1%浓度的花多多1号水溶肥(氮磷钾比例为20∶20∶20),保证大豆苗期水肥供应充足. 所有大豆幼苗在T1L1处理下生长,待第一片三出复叶完全展开且第二片复叶刚露尖时,将部分幼苗分别移至其他3个处理下. 隔5 d调查株高、茎粗、节间长动态,在V3、V5时期取样测定大豆叶面积和干物质积累,并取倒二功能叶测定光合特性.
-
将大豆幼苗置于不同处理条件下,每个处理选取5株生长状况相近的大豆植株,调查其株高、茎粗和节间长的动态变化,每次调查间隔5 d,共5次,重复3次.
-
大豆V3和V5期时,每个处理挑选3株长势一致的大豆幼苗,将每株所有叶片摘下,平整且不重叠地放置在黑布上,同时放上标尺和编号牌,对其进行拍照,用Image-Pro Plus 6.0软件计算叶面积,重复3次.
-
大豆V3和V5期时,每个处理选取3株长势一致的大豆幼苗,称取茎、叶的鲜质量,装进牛皮纸袋中于烘箱105 ℃杀青1 h,80 ℃烘干至恒重,再称量并记录干重,重复3次.
-
大豆V3和V5期时,每个处理选取3株长势一致的大豆幼苗,使用LI-6800高级光合荧光测量系统于当日08:00至12:00对其倒二功能叶中间叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间二氧化碳浓度(Ci)进行测定,设定风扇转速(Fan Speed)=1 000 r/min,流量=500 μmol·mol-1,CO2浓度=400 μmol·mol-2,重复3次.
-
大豆V3和V5期时,每个处理挑选3株长势一致的大豆幼苗,使用LI-6800高级光合荧光测量系统和叶绿素荧光成像系统CFI对其倒二功能叶中间叶片的荧光参数进行测定,测定指标包括大豆叶片的光系统II(PSII)最大光化学量子产量(Fv/Fm)、光化学荧光猝灭系数(qP)、PSII实际光化学效率(Fq′/Fm′)和非光化学荧光猝灭系数(NPQ). 测定过程中,使大豆叶片充分暗处理15 min稳定后获得暗反应数据,开启光照设置光强为1 000 μmol/(m2· s),光处理叶片15 min稳定后获得光处理数据,重复3次.
-
大豆V3和V5期时,每个处理挑选3株长势一致的大豆幼苗,剪碎其倒二功能叶,称取0.5 g放入装有丙酮∶乙醇(体积比1∶1)10 mL溶液的试管中,置于恒温箱25 ℃黑暗环境下浸提48 h,并每隔数小时对其进行震荡,浸提至叶片成乳白色后,对浸提液进行过滤,之后置于663 nm、645 nm、470 nm波长下测定上清液的吸光度[10-12]得出总叶绿素含量、叶绿素a/b以及类胡萝卜素的数值,重复3次.
其中:A663、A645、A470分别代表波长663 nm、645 nm和470 nm所得的吸光度值. V为稀释后的溶液体积,单位为mL. W为称取的大豆叶片鲜质量,单位为g.
-
试验数据采用Microsoft Excel 2021进行汇总整理,用OriginLab Origin 2022进行数据分析与作图.
1.1. 试验材料
1.2. 试验设计
1.3. 测定项目与方法
1.3.1. 生长动态
1.3.2. 叶面积
1.3.3. 干物质积累
1.3.4. 光合参数
1.3.5. 叶绿素荧光参数
1.3.6. 叶绿素含量
1.4. 数据处理与分析
-
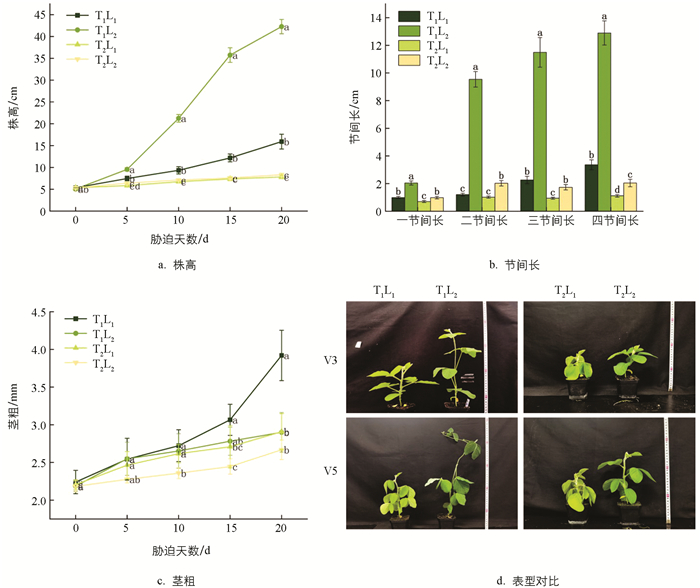
低温和荫蔽胁迫显著改变大豆的表型(图 1). 荫蔽胁迫会使大豆幼苗株高和节间长徒增,在胁迫第20 d时,T1L2(荫蔽)处理的株高较T1L1(正常温度正常光,简称CK)处理增加了165.68%,但会抑制茎粗,较T1L1(CK)处理降低了26.02%. 低温胁迫显著抑制大豆幼苗的株高、茎粗和节间长,在胁迫第20 d时,T2L1(低温)处理的株高和茎粗较T1L1(CK)处理分别降低了50.91%和25.77%. 低温和荫蔽双重胁迫显著抑制大豆幼苗株高和茎粗,但由于荫蔽的影响,对大豆节间长的抑制不明显,且相比于低温单胁迫,株高和节间长依旧徒增,在胁迫第20 d时,T2L2(低温和荫蔽)处理的株高和茎粗较T1L1(CK)处理降低了47.64%和31.89%.
-
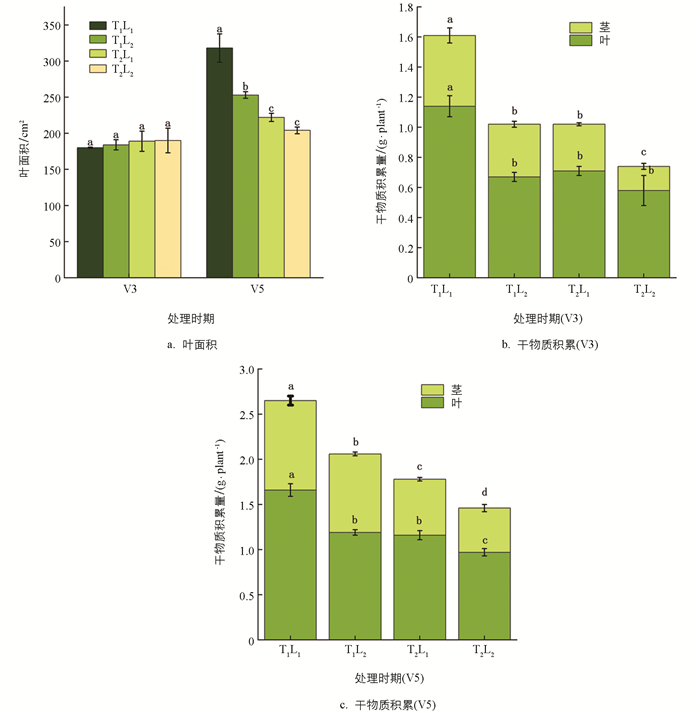
低温和荫蔽胁迫抑制大豆的光合作用,从而抑制叶面积的增加和干物质的积累(图 2). 荫蔽胁迫抑制大豆幼苗叶面积和干物质积累,V5时期,T1L2(荫蔽)处理的叶面积和干物质积累量较T1L1(CK)处理降低了20.44%和22.26%. 低温胁迫也抑制大豆幼苗叶面积和干物质积累,V5时期,T2L1(低温)处理的叶面积和干物质积累量较T1L1(CK)处理降低了30.19%和32.83%. 低温和荫蔽双重胁迫显著抑制大豆幼苗叶面积和干物质积累,在V5时期,T2L2(低温和荫蔽)处理的叶面积和干物质积累量较T1L1(CK)处理降低了35.85%和45.28%.
-
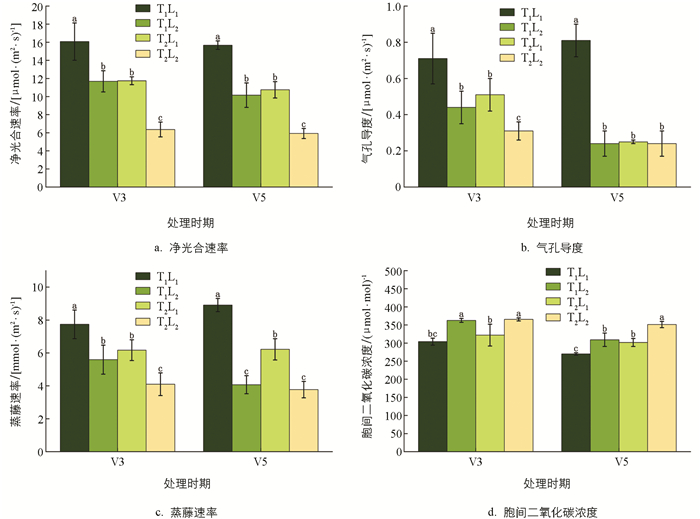
低温和荫蔽会抑制大豆幼苗的光合速率,使得Pn、Gs和Tr下降,Ci上升(图 3). 荫蔽胁迫下,Pn、Gs和Tr显著下降,Ci显著上升,V5时期,T1L2(荫蔽)处理的Pn、Gs和Tr较T1L1(CK)处理分别降低35.27%、70.37%和54.32%,Ci上升14.49%. 低温胁迫下,Pn、Gs和Tr显著下降,Ci显著上升,V5时期,T2L1(低温)处理的Pn、Gs和Tr较T1L1(CK)处理分别降低35.59%、69.14%和30.19%,Ci上升11.76%. 低温和荫蔽双重胁迫下,大豆幼苗的光合参数值较单胁迫变化更明显,对光合速率的抑制程度更严重,V5时期,T2L2(低温和荫蔽)处理的Pn、Gs和Tr较T1L1(CK)处理分别降低60.33%、70.37%和57.69%,Ci上升30.08%.
-
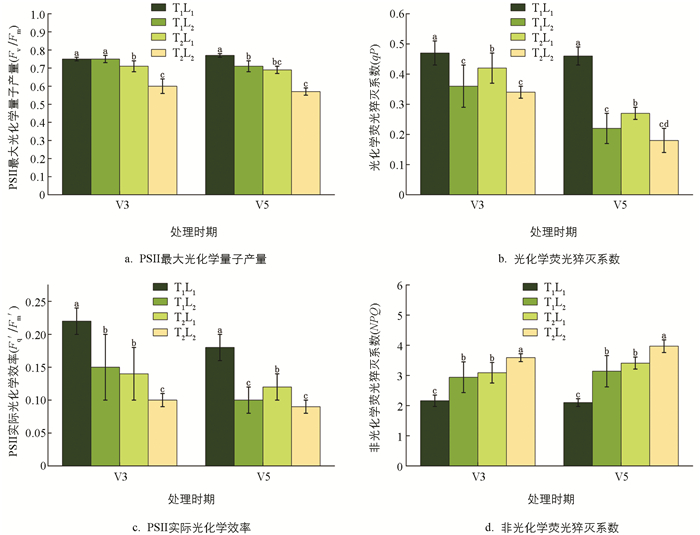
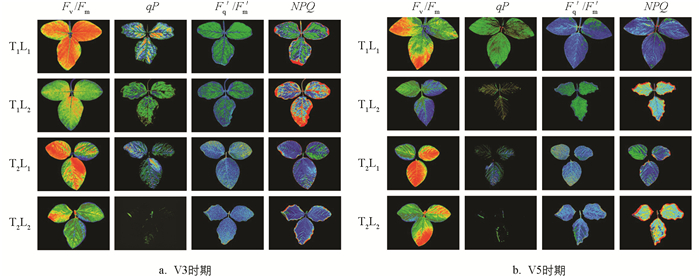
低温和荫蔽胁迫下大豆幼苗的Fv/Fm、qP和Fq′/Fm′呈下降趋势,NPQ呈上升趋势(图 4、图 5). V5时期,T1L2(荫蔽)处理的Fv/Fm、qP和Fq′/Fm′较T1L1(CK)处理分别降低7.79%、52.17%和44.44%,NPQ上升49.52%. T2L1(低温)处理的Fv/Fm、qP和Fq′/Fm′较T1L1(CK)处理分别降低10.39%、41.30%和33.33%,NPQ上升62.38%. T2L2(低温和荫蔽)处理的Fv/Fm、qP和Fq′/Fm′较T1L1(CK)处理分别下降了25.97%、60.87%和50.00%,较T1L2处理下降了19.72%、31.82%和10.00%,较T2L1处理下降了17.39%、33.33%和25.00%;NPQ较T1L1(CK)处理上升了89.05%,较T1L2处理上升了26.43%,较T2L1处理上升了16.42%.
-
由表 2可知,V5时期,T1L2(荫蔽)处理的Chla、Chlb、Car和Chla/b较T1L1(CK)处理分别上升了67.82%、37.84%、17.39%和18.67%,其中,荫蔽胁迫对Car影响显著,对Chla、Chlb和Chla/b的影响极显著. T2L1(低温)处理Chla和Chlb较T1L1(CK)处理下降了13.79%和24.32%,Car和Chla/b上升了4.35%和12.45%,其中,低温胁迫对Chla/b的影响显著,对Chla、Chlb和Car的影响极显著. T2L2(低温和荫蔽)处理Chla、Chlb和Car较T1L1(CK)处理分别降低20.69%、37.84%和13.04%,Chla/b上升25.73%,其中,低温和荫蔽双重胁迫对Chla、Chlb和Car的影响极显著.
2.1. 低温和荫蔽双重胁迫对大豆苗期生长的影响
2.1.1. 株高、茎粗和节间长
2.1.2. 叶面积和干物质积累
2.2. 低温和荫蔽双重胁迫对大豆苗期光合特性的影响
2.2.1. 光合参数
2.2.2. 叶绿素荧光参数
2.2.3. 叶绿素含量
-
遭受低温冷害的大豆生育期延后,生育日数明显增加[12],其株高、主茎节数、叶片鲜质量、茎鲜质量都显著降低[6]. 荫蔽胁迫下的大豆植株茎秆纤细、陡长、易倒伏[13],总生物量和叶面积显著降低[14]. 在本试验中,低温和荫蔽双重胁迫处理下,大豆的生育期延长,株高、茎粗显著降低,二节间长显著升高,叶面积和干物质积累随着时间的延长下降趋势明显. 可能是由于大豆在低温和荫蔽的双重胁迫下,地上部分光合作用受到的阻碍更加严重,导致大豆的生长受到严重伤害,且相比于低温和荫蔽单一胁迫处理所受到的伤害更严重.
-
影响光合速率降低的因素有两种,分别是气孔限制因素和非气孔限制因素,当Ci随着Pn、Gs及Tr的下降而下降,则气孔因素占主导[15];当Ci随着Pn、Gs及Tr下降而升高,则非气孔因素占主导[16]. 本试验中,低温和荫蔽双重胁迫处理下,大豆叶片的Pn、Gs、Tr显著降低,Ci显著升高,且相较于低温和荫蔽单一胁迫处理,变化幅度更大. 可能是由于双重非气孔因素(低温和荫蔽)对大豆光合速率的抑制作用更严重.
叶绿素荧光参数反映植物在胁迫条件下,光能的分配、耗散及光合机构的损伤[17]. 逆境环境下,Fv/Fm降低表示PSII发生了光抑制[18],光化学猝灭系数qP是光化学反应所引起的荧光产额的下降,反映PSⅡ原初电子受体(QT)的还原状态及开放的反应中心占总PSⅡ反应中心的比例[19]. 本试验中,低温和荫蔽双重胁迫处理下,大豆叶片的Fv/Fm、qP和Fq′/Fm′均显著降低,可能是由于大豆叶片的PSⅡ功能下降,发生光抑制,且相较于低温和荫蔽单一胁迫处理,Fv/Fm、qP和Fq′/Fm′降低更显著,说明在低温和荫蔽双重胁迫下,光抑制程度可能更明显. NPQ是指PSⅡ天线色素吸收的光能不能用于光合电子传递而以热能的形式耗散掉的光能部分,对光合系统起一定的保护作用[20],可用来衡量植物对光系统损害的抵御程度,NPQ值越高,则对外界的不良光系统损害抵抗力越强[21]. 本试验中,低温和荫蔽双重胁迫处理下,大豆叶片的NPQ显著升高,说明大豆在受到低温和荫蔽双重胁迫时可能开启自我保护机制,通过NPQ的升高来抵御外界不良光系统.
光合色素包括叶绿素和类胡萝卜素[22],具有吸收、传递和转化光能的作用. 叶绿素含量常作为叶绿体发育、光合能力的指标,与光合速率呈正相关[23],类胡萝卜素可以起到光能捕获和光破坏防御的作用[24]. 本试验中,低温和荫蔽双重胁迫处理下,大豆叶片的Chla、Chlb和Car均显著降低,且均低于低温和荫蔽单一胁迫处理,可能是由于在低温和荫蔽双重胁迫下,对大豆叶片中叶绿素合成酶活性的抑制作用更显著,导致叶绿素含量降低,影响大豆幼苗的光合作用.
3.1. 低温和荫蔽双重胁迫对大豆苗期生长的影响
3.2. 低温和荫蔽双重胁迫对大豆苗期光合特性的影响
-
综上,与正常温度正常光、正常温度荫蔽和低温正常光相比,低温和荫蔽双重胁迫下,大豆的株高、茎粗、叶面积和干物质积累量显著降低. 净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)显著降低,胞间二氧化碳浓度(Ci)显著升高;PSII最大光化学量子产量(Fv/Fm)、光化学荧光猝灭系数(qP)和PSII实际光化学效率(Fq′/Fm′)显著降低,非光化学荧光猝灭系数(NPQ)显著升高;叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)显著降低,叶绿素a/b(Chla/b)显著升高. 说明低温和荫蔽双重胁迫会抑制大豆的生长和光合特性,且相比于低温或荫蔽单一胁迫下,抑制作用更加明显.