



 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
葡萄(Vitis vinifera L.)原产于亚洲西部和欧洲,距今已有5 000多年栽培历史;我国栽培葡萄是从西汉开始,至今也已有2 000多年,是我国和世界上种植面积最广泛和深受世人衷爱的一种水果[1-3]. 葡萄果实含有人体所需的氨基酸、葡萄糖等糖类、磷钾钙铁锰镁等矿质元素及多种维生素(Vb1,Vb2,Vb6,Vb12,Vp),具有抗氧化、保护心血管、抗病毒、保肝及抗癌等功效,美味营养,适量食用可以健胃消食,促进人体健康[4-6]. 另外,葡萄的经济价值也很高,种植葡萄是云南贫困山区脱贫致富的一条重要途径.
植物病害是影响葡萄生长发育、产量及品质的一类重要生物灾害因子,国内外已有报道的葡萄病害达40多种,其中主要有葡萄霜霉病(Plasmopara viticola)、白粉病(Uncinula necator)、黑痘病(Sphaceloma ampelinum)、灰霉病(Botrytis cinneria)、褐斑病(Pseudocercospora vitis)及轮斑病(Acrospermun viticola)等真菌病害[7-9],而炭疽病则是在葡萄产区发生流行和为害严重的病害之一,其由不同的刺盘孢种类(Colletotrichum spp.)真菌侵染所致,国际上已报道的有盘长孢刺盘孢(C. gloeosporioides)、尖孢刺盘孢(C. acutatum)、果生刺盘孢(C. fructicola)、葡萄刺盘孢(C. viniferum)和隐秘刺盘孢(C. aenigma)等[10-11],我国各省(区、市)已报道的有盘长孢刺盘孢、葡萄刺盘孢、隐秘刺盘孢及河北刺盘孢(C. hebeiense)[6, 7, 12-16],但不同国家和地区所报道的葡萄炭疽病病原菌种类存在较大的差异.
云南是我国重要的葡萄产区之一,2021年葡萄种植面积和产量分别为39 000 hm2和975 000 t[17],炭疽病在该地区葡萄上全域发生,几乎连年流行,造成严重的产量和经济损失. 2013年邓维萍等[18]从云南东部几个县(市)采样鉴定,葡萄炭疽病的病原菌为盘长孢刺盘孢(C. gloeosporioides)和尖孢刺盘孢(C. acutatum),而对于滇西地区发生的葡萄炭疽病,文献中尚无病原菌鉴定研究的有关报道. 为此,本研究于2022-2023年从滇西不同葡萄种植区采集病样进行分离纯化病原菌,应用多基因分子生物学方法进行鉴定,在此基础上筛选出高效低毒的杀菌剂,为该地区葡萄炭疽病的诊断监测和综合防控提供科学依据.
全文HTML
-
本研究于2022-2023年在云南西部的迪庆、保山、临沧和普洱等不同地区的葡萄园采集具有典型炭疽病症状的病叶样品26份,带回室内供病原菌分离纯化.
-
采用北京三药科技开发公司生产的马铃薯葡萄糖琼脂(PDA)培养基,使用时在1 000 mL蒸馏水中加37 g原药粉,在电磁炉上煮沸5 min,边加热边搅拌,至原药粉完全均匀分散于水中成酱汁后分装于250 mL锥形瓶中,瓶口盖上封口膜蘑后放入高压灭菌锅中灭菌,在温度121 ℃ (0.10~0.12 Mpa)下灭菌30 min后取出,待冷却至50~60 ℃时倒成培养基平板,分别用于病原菌分离纯化及杀菌剂抑菌测定.
-
本研究参考有关文献选取扩增病原菌的核糖体内转录间区(ITS)、肌动蛋白、钙调蛋白、组蛋白3和微管蛋白基因(ACT,CAL,HIS3和TUB2)序列,所用引物对的详情见表 1,它们均由上海生物工程有限公司合成.
-
选择当前生产中广泛应用于防治作物真菌性病害的8种杀菌剂,用于药剂筛选试验,主要包括80%代森锰锌WP、64%噁霜·锰锌WP、12.5%百菌清WP、30%肟菌·戊唑醇SA、12.5%四氟醚唑WP、40%苯醚甲环唑SA、25%甲硫已唑醇SA和30%苯甲吡唑脂SA,均由云南景洪市楚杰农资有限责任公司提供.
-
于2022-2023年在葡萄生长季节(夏秋季)到迪庆、保山、临沧和普洱葡萄园观察(拍照)记录炭疽病的发生情况,并与后续实验室接种试验形成的病害症状进行比较. 在调查时采集具有典型炭疽病症状的病叶,带回实验室做病原菌分离纯化.
-
将采集的新鲜病叶用自来水清洗干净并晾干后再用病组织分离法[20]分离纯化真菌. 在病斑与周围健康组织交界位置切取约5 mm×5 mm的病斑组织小块,放置于75%乙醇液体中30 s,移入2%的次氯酸钠溶液中2 min,依次在灭菌蒸馏水中清洗3次,最后置于灭菌滤纸上吸干组织块表面残留的水分. 在超净台上用消毒灭菌的镊子将组织块植入培养皿中的PDA平板上,放入(25±2) ℃黑暗恒温箱中培养. 待菌落形成后用灭菌接种针挑取边缘菌丝体尖端,植入新的PDA平板中央,在恒温箱中培养至形成菌落;再挑取菌丝尖端纯化2次而得到纯培养真菌菌株,最后从无污染的菌落边缘挑取菌丝尖端植入试管PDA斜面上,在恒温箱中生长4 d后放入4 ℃冰箱中保存备用.
-
采用微刺伤离体叶片接种法[20]. 从温室栽植的健康葡萄(品种为巨峰)植株上采集健康叶片,用自来水冲洗干净后置于直径15 cm培养皿中的湿滤纸片上;用灭菌缝衣针针尖在接种点位处轻轻刺伤叶片表面,用灭菌打孔器(直径5 mm)从培养7 d的各菌株菌落边缘取菌饼,置于刺伤点位处. 在每个叶片接种4个位点,每个菌株重复3个叶片,以叶片不接种作为对照. 处理后用灭菌水做叶面喷雾后盖上培养皿,在室温下黑暗保湿(RH=100%)24 h,然后置入生长培养箱中培养至14 d;培养条件:(25±2) ℃,RH>90%,11 000 lux光照12 h/d;定期观察发病情况. 接种36 h后移去菌饼,10 d后取发病叶片的病斑组织再分离纯化其病原菌,并与原来菌株形态进行比较,根据柯赫氏病证法则[21]确认致病菌株.
-
从PDA平板培养基上培养7 d的病菌菌落上刮取菌体,用溴化十六烷基三甲基铵(CTAB)法[22]提取基因组DNA,进行不同基因序列的扩增. PCR反应液(25 μL)配方为:2×EasyTaq PCR Super Mix 12 μL、引物各1 μmol/L、模板DNA 1 μL、重蒸水10 μL.
① 分别用ITS1/ITS4和ACT512F/ACT783R引物对扩增内转录间区(internal transcribed spacer,ITS)序列和肌动蛋白基因(actin gene,ACT);扩增程序设置为:95 ℃退火3 min,(95 ℃退火60 s、52 ℃解链30 s和72 ℃延伸1 min) 循环34次,最后72 ℃延伸10 min[23].
② 用CL1/CL2引物对扩增钙调蛋白基因(Calmodulin gene,CAL);扩增程序设置为94 ℃变性2.5 min,(94 ℃变性30 s,50 ℃退火30 s,72 ℃延伸30 s) 34次循环,最后72 ℃延伸10 min[24].
③ 用CYLH3F/CYLH3R引物对扩增组蛋白3基因(histone 3 gene,HIS3);扩增程序设置为:96 ℃变性5 min,(96 ℃变性30 s、52 ℃退火30 s和72 ℃延伸1 min)循环34次,最后72 ℃延伸5 min[25].
④ 用T1/Bt2b引物对扩增微管蛋白基因(β-Tubulin gene,TUB2);扩增程序设置为:94 ℃变性4 min,(94 ℃变性45 s、60 ℃退货45 s和72 ℃延伸1 min) 循环34次,最后72 ℃延伸10 min[24].
各种PCR产物送上海生物工程有限公司分别回收和测序. 将获得的各病原菌的基因序列上传到NCBI GenBank数据库,在线(
https://blast.ncbi.nlm.nih.gov/Blast.cgi ) 进行Blast-n对比分析;用DNAman软件将获得的各菌株蛋白基因序列进行拼接,再用MEGA7.0[26]构建以拼接序列为基础的邻接(neighbor-joining)系统演化树分析;根据分析结果确定病原菌的物种名称. -
将已鉴定的不同种病原菌接种到PDA平板培养基上,在(25±2) ℃恒温箱中黑暗培养7~10 d后观察记录菌落形态,14~20 d后在显微镜下观察记录分生孢子特征并在显微镜(40倍)下测定30根菌丝和30个分生孢子的大小(长×宽);将分生孢子悬浮涂抹液接种到贴于载玻片上的洋葱鳞片内表皮膜上,在(25±2) ℃恒温箱中保湿培养48~72 h后在显微镜下观察附着胞的形态,测定30个附着胞的大小(长×宽).
-
选取优势病原菌(C. gloeosporioides PT23-01),分别进行药剂种类筛选、最有效药剂的毒力分析和控病作用试验.
① 药剂筛选.用菌落生长法[27]测试8种当时被广泛应用的杀菌剂作为供试药剂. 预先在PDA上培养鉴定的优势病原菌(C. gloeosporioides PT23-01),按生产商推荐的使用浓度中值作为测试浓度,在超净工作台上用灭菌水配制药液;用移液枪取200 μL每种配制好的药液置入PDA平板培养基表面,用灭菌玻璃涂抹棒涂抹使药液均匀布满培养基表面,以同体积的无菌水涂抹DNA平板作为对照. 用灭菌打孔器从PDA培养的致病菌菌落边缘取直径5 mm的菌饼,放置于培养基平板中央;每个药剂重复处理5皿. 处理后放入(25±2) ℃恒温箱中培养6 d后测量菌落直径(cm),计算其平均值及标准误,用邓肯氏新复极差法[28]检验不同处理的菌落直径值间的差异显著性.
根据直径值计算出各种杀菌剂的抑菌率=(对照菌落直径-药剂处理菌落直径)/对照菌落直径×100%.
② 药剂对不同病菌的抑菌作用测试. 选择上述试验中对PR23-01抑菌作用不同(最弱、居中、最强)的3种药剂(80%代森锰锌WP、12.5%四氟醚唑WP、30%苯甲·吡唑脂SA),按生产商推荐的使用浓度中值作为测试浓度,测试它们对3种不同种刺盘孢菌的抑菌作用.
③ 药剂的毒力分析.选取抑菌率最高的2种药剂(25%甲硫乙唑醇SA和30%苯甲吡唑脂SA),分别用不同的浓度梯度的药剂测试抑菌率,按照李建明等的方法[29]进行毒力分析,构建毒力回归直线模型,用F测验法[28]检验模型的显著性,并通过建立的模型计算各药剂的有效中浓度值(EC50).
④ 药剂的控病效果测试.选用抑菌效果不同的4种药剂进行室内植物药剂控病试验. 用PDA在(25±2) ℃培养14 d的PTTJ01,10 d后从菌落上收集分生孢子和菌丝混合物,用灭菌水配制成菌体悬浮菌液(约105个/mL),喷雾接种于温室内盆栽4个月的健康葡萄(品种为巨峰)苗上,至其叶片表面均匀布满菌液,每种药剂重复5个植株(钵);用灭菌水喷雾5钵植株作为对照;将接种苗转入人工气候箱中,在25 ℃、RH 100%和黑暗条件下培植24 h,然后在25 ℃,RH>90%,11 000 lux光照10 h/d下继续培植48 h;用配制的不同杀菌剂药液喷雾,以清水喷雾植株作为对照. 喷药处理后继续培植21 d,期间适时观察记录植株发病情况,每个处理观察20片功能叶,记录每个叶片的病害严重度(病斑面积占总叶面积的百分比)并计算平均严重度,用邓肯氏新复极差法[28]检验不同处理的差异显著性,并计算各药剂的控病效果.
1.1. 供试材料
1.1.1. 葡萄病叶样品
1.1.2. 培养基
1.1.3. 引物
1.1.4. 杀菌剂
1.2. 研究方法
1.2.1. 葡萄病叶样品采集
1.2.2. 真菌的分离纯化
1.2.3. 致病性试验
1.2.4. 病菌的PCR分子鉴定
1.2.5. 病原菌形态特征观察
1.2.6. 杀菌剂筛选
-
用PDA平板培养基分离和纯化,从葡萄病叶病组织样品获得13个菌体形态明显不同的纯培养菌株. 通过离体叶片微刺伤接种后,有7个菌株(命名为PT23-01~PT23-07)引起侵染发病,接种48 h后在菌饼下的叶组织软化呈水渍状,在接种5~7 d后各菌株形成非常相似的病斑(图 1a,b),其成圆形或椭圆形,表面和背面均褐色,病斑外有黄色晕圈. 取接种后7 d的病斑组织再进行分离纯化,获得了菌落形态与7个菌株相同的纯培养. 用其他6个菌株处理21 d后也未见叶片发病,因此认为它们不是致病菌. 未接种的对照叶片一直保持健康状态,一直未见发病(图 1c). 由此试验结果并根据柯赫氏证病法则确认,PT23-01~PT23-07是引起滇西地区葡萄炭疽病的病原菌.
-
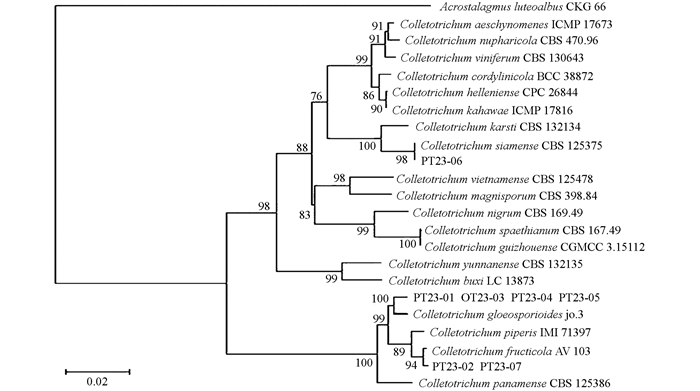
用CTAB方法分别从PT23-01~PT23-07菌株培养物中提取其基因组DNA,通过用不同的引物进行PCR扩增,获得它们的ITS以及ACT,CAL,HIS3和TUB2基因序列,将这些序列上传到NCBI GenBank数据库,在线进行Blast-n比对分析. 结果表明,PT23-01,PT23-03,PT23-04和PT23-05菌株与盘长孢刺盘孢(Colletotrichum gloeosporioides)相应菌株的相似百分率(per. identity)为99.27%~100%;菌株PT23-02和PT23-07与果生刺盘孢(C. fruticola)相似率为99.69%和100%;而PT23-06菌株则与暹罗刺盘孢(C. siemense)相似率为100%.
参考Blsat-n分析结果,从GenBank数据库中下载不同Colletotrichum spp.和白黄笋顶孢霉(Acrostalagmusluteoalbus)CKG66菌株(外群)的相应蛋白基因序列,将7个菌株及下载菌株的4个蛋白基因序列分别拼接而获得它们的拼接序列. 用MAGA7.0构建基于各物种菌株基因拼接序列的系统演化树(图 2). 从该系统演化树可看出,PT23-01,PT23-03,PT23-04和PT23-05菌株与盘长孢刺盘孢(Colletotrichum gloeosporioides) jo-3菌株位于同一末端分枝上,Bootstrap支持率BJ=100%;PT23-02和PT23-07与果生刺盘孢(C. fruticola) AV103菌株位于另一末端分枝上,BJ=94%;而PT23-06则与暹罗刺盘孢(C. siemense) CBS125375菌株处于同一末端分枝,BJ=98%. 系统演化分析的结果与Blast-n比对结果完全一致.
-
在不同生长季节调查自然葡萄园,结果发现,炭疽病在葡萄的各个生长发育阶段均可侵染. 在苗床的幼苗叶片上的病斑呈圆形或不规则形,中部褐色(图 3a). 在田间或葡萄园结果植株上,藤蔓尖端叶片与幼苗上的病斑相似,多为圆形或椭圆形,少有呈不规则形,略凹陷;病斑直径多为4.2~13.7 mm,中央区呈褐色,周围褐色环多不明显,黄色晕圈明显(图 3b). 生长季节早期叶片上的病斑较规则,圆形或椭圆形,褐色中央区域被一浅褐色带包围,浅褐色带外再围绕一较齐整的褐色环,病斑外偶尔可见黄色晕圈(图 3c). 生长季晚期叶片上的病斑大小差异较大,多为不规则形(图 3d,e),可扩展或联合形成不同大小的枯斑,从而导致叶片大面积黄化枯死(图 3f).
-
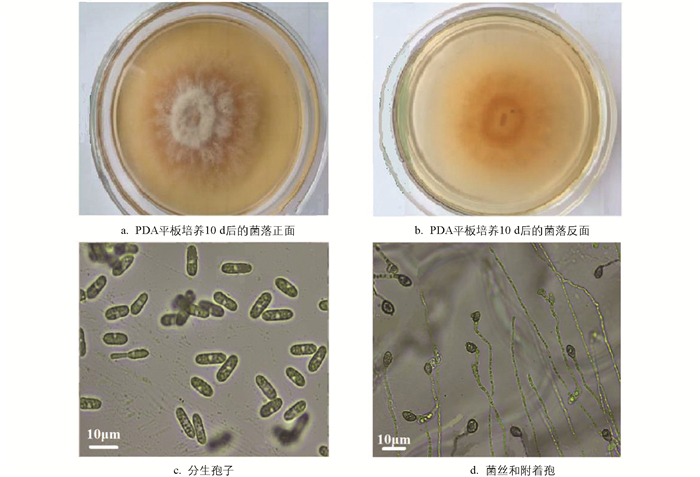
盘长孢刺盘孢在PDA平板培养基上的形态特征如图 4所示. 在(25±2) ℃条件下培养5 d后形成的菌落呈圆形或近圆形,正面白色,背面中间区域橘红色,边缘白色;气生菌丝较发达,有轮纹,直径(55.68±3.28) mm (n=5),生长速率11.16 mm/d;培养10 d后在菌落的菌丝层下形成橙色的分生孢子团;在显微镜下观察,菌丝无色,分枝较发达,分隔,直径1.83~4.26 (3.25) μm (n=50). 分生孢子梗分化不明显;分生孢子无色,单细胞(不分格),直或稍弯曲,长椭圆形,两端盾圆,11.82~17.41 (14.62) μm×4.15~5.48 (4.86) μm (n=50). 分生孢子在洋葱鳞片表皮膜上萌发24 h后形成不同长度的芽管,48 h后可见较多附着孢,深褐色,大多圆形或梨形,少有不规则形,5.21~7.65 (6.62) μm×4.53~5.78 (5.64) μm (n=50). 病菌的这些特征基本符合Weir等[24]、Hassan等[30]和Rabha等[31]记述的盘长孢刺盘孢(Colletotrichum gloeosporioides Penz.)的形态学特征.
-
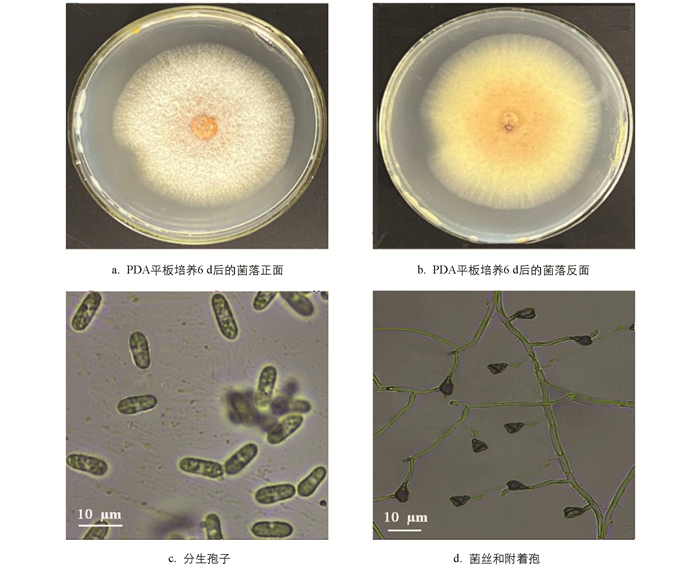
在(25±2) ℃恒温箱中培养6 d后,菌落圆形或椭圆形,初期呈白色或粉白色,表面绒毡状,边缘较整齐,背面略呈桔红色,直径(65.86±6.31) mm (n=5),生长速率(10.98±1.02) mm/d;后期随着大量分生孢子团的产生而使菌落表面部分变成红色. 在显微镜下观察,该菌的菌丝透明,分隔,直径1.28~2.63 (2.07) μm (n=50). 分生孢子无色,单胞,椭圆或长椭圆形,两端盾圆或稍尖细,细胞内有1~3个油滴,9.3~18.27 (14.1) μm×4.3~6.2 (5.2) μm. 附着胞黑褐色,单细胞,椭圆形或三角形,4.82~7.75 (6.31)×4.23~5.92 (5.15) μm (n=50)(图 5). 所述这些形态特征与Lu等[32]描述的果生刺盘孢(Colletotrichum fructicola Prihast.,L. Cai & K.D. Hyde)吻合.
-
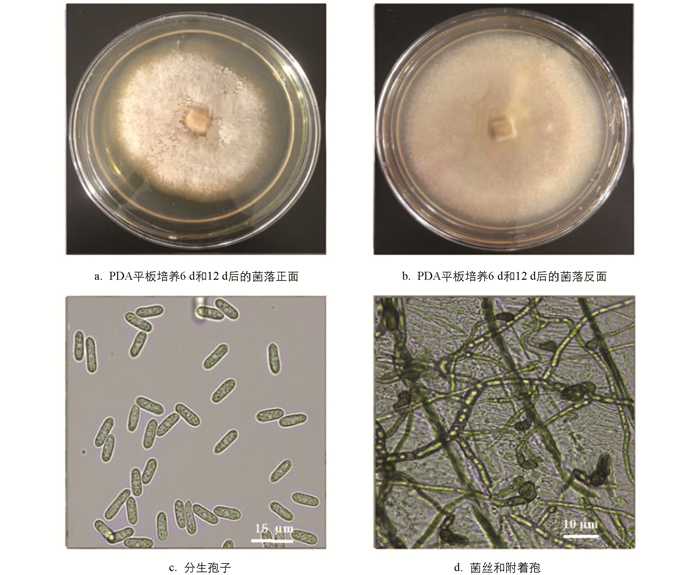
在PDA平板上(25±2) ℃培养6 d后,其菌落圆形,表面灰白色,棉絮状,直径(69.6±5.2) mm (n=5),生长速率(11.6±0.9) mm/d. 菌丝浅色或无色,分隔,呈近直角分枝,直径1.25~3.07 μm (n=50);培养12 d后菌落灰色加深,表面略呈粉状,在菌落上形成分生孢子,无色,单细胞不分隔,长椭圆形,两端盾圆或一端梢尖斜,9.38~16.81 (13.62)×4.20~6.27 (5.26) μm (n=50). 分生孢子在洋葱表皮上萌发形成菌丝,顶端产生附着胞,褐色或深褐色,椭圆形或少不规则形,5.9~9.3 (7.6)×3.8~6.4 (4.8) μm (n=50) (图 6). 这些特征与李维峰等[20]和Weir等[24]描述的C. siamense特征一致.
-
试验结果表明,供试的8种杀菌剂对盘长孢磁盘孢菌(PT23-01)都具有显著的抑制作用(表 2). 在设定的药剂浓度下,病原菌菌落生长受到一定程度的抑制,其中64% 噁霜·锰锌WP、80%代森锰锌WP、75%百菌清WP和30%肟菌·戊唑醇SA的抑菌作用较弱,抑菌率<40%;12.5%四氟醚唑WP和40%苯醚·甲环唑SA的抑菌作用较强,抑菌率<90%;25%甲硫·乙唑醇SA和30%苯甲吡唑脂SA的抑菌效果最好,抑菌率分别为92.25%和95.87%.
-
本研究测定了不同的3种杀菌剂对本研究鉴定的葡萄3种刺盘孢的抑制作用,结果显示,同一个药剂对不同种刺盘孢的抑制作用非常一致. 80%代森锰锌WP对3种不同刺盘孢的抑菌作用效果较差,抑菌率只有50%左右;12.5%四氟醚唑WP的抑菌效果居中,抑菌率为76.67%~78.28%;而30%苯甲吡唑脂SA的抑菌效果最好,抑菌率达90.37%~93.25%. 另外,同一种药剂对3种不同刺盘孢菌的抑菌作用没有显著差异(表 3).
-
选择在生产商推荐使用浓度下对PT23-01抑菌效果最好的25%甲硫乙唑醇SA和30%苯甲吡唑脂SA两种药剂,分别设置适当的浓度,进一步测试它们在不同浓度下的抑菌作用. 试验结果表明,药剂的抑菌率和药剂浓度呈显著的正相关关系,根据测定结果建立两种药剂的毒力回归方程分别为:
方程中x和y分别表示杀菌剂浓度(mg/mL)和抑菌率(%),经F检验两个方程都达到极显著水平(p<0.001);由毒力回归方程分别计算出它们的有效抑菌中浓度(EC50)分别为1.103 mg/mL和1.521 mg/mL. 由此可见,两种杀菌剂对病菌的有效抑制中浓度都很低,表明它们的抑菌效果都非常强.
-
温室盆栽葡萄苗上的药剂控病试验结果显示,处理21 d后未用药的对照葡萄植株炭疽病的严重度为25.52%. 供试的4种杀菌剂处理都显著(p<0.01)降低了葡萄炭疽病的严重度,但不同药剂处理植株的病害严重度之间也存在着显著差异,因此它们对炭疽病的控制作用也很不相同;其中12.5%四氟醚唑WP和40%苯醚·甲环唑SA的防病效果不够理想,控病率只有36.40%和55.52%;而25%甲硫·乙唑醇SA和30%苯甲·吡唑脂SA的防病效果则很好,其控病率分别达85.03%和91.54%,用它们处理后葡萄炭疽病严重度分别降低到3.82%和2.16%,植物整体上保持健康,只有个别叶片上可见少量的小病斑(表 4).
2.1. 病原菌的分离纯化与致病性测定
2.2. 病原菌的分子鉴定
2.3. 葡萄炭疽病的症状
2.4. 病原菌的形态学特征
2.4.1. 盘长孢刺盘孢(C. gloeosporioides:PT23-01,PT23-03,PT23-04和PT23-05)
2.4.2. 果生刺盘孢(C. fruticola:PT23-02和PT23-05)
2.4.3. 暹罗刺盘孢(C. siemense:PT23-06)
2.5. 杀菌剂的抑菌和控病作用
2.5.1. 不同杀菌剂的抑菌作用
2.5.2. 不同杀菌剂的抑菌效果
2.5.3. 两种药剂的毒力分析
2.5.4. 药剂的控病效果
-
由刺盘孢属真菌(Colletotrichum spp.)侵染所致的炭疽病在全球葡萄种植国家和地区常年发生和危害,是葡萄上的重要流行性病害之一. 据Kim等[10]及Suzaki[11]报道,日本、澳大利亚、韩国和加拿大等国家发生的主要有盘长孢刺盘孢(C. gloeosporioides)、尖孢刺盘孢(C. acutatum)、果生刺盘孢(C. fructicola)、葡萄刺盘孢(C. viniferum)和隐秘刺盘孢(C. aenigma)等;我国各省(区、市)已报道的葡萄炭疽病病菌则有盘长孢刺盘孢、葡萄刺盘孢、隐秘刺盘孢及河北刺盘孢(C. hebeiense)[6-7, 13-15],有的地方同时记载几种病菌侵染. 2013年邓维萍等[18]从云南东部地区采集的样品中鉴定出盘长孢刺盘孢和尖孢刺盘孢两个种. 本研究结果表明,云南西部地区发生的葡萄炭疽病病原菌有盘长孢刺盘孢(C. gloeosporioides)、果生刺盘孢(C. fructicola)和暹罗刺盘孢(C. siamense),而未发现其他已报到的真菌物种. 在滇西地区3种葡萄炭疽病菌中,盘长孢刺盘孢在所有调查的地区均有发生分布,是优势病原菌种群;果生刺盘孢在迪庆和临沧地区有发生;而暹罗刺盘孢只在普洱地区被发现,是作者鉴定出的一种葡萄炭疽病新病原菌. 因此,在滇西地区葡萄炭疽病的防治,应以盘长孢刺盘孢为主要防治对象物种,同时兼顾另外两种病菌,根据它们的生物学特性及其所致炭疽病的发生流行规律制订经济有效的综合防控技术.
刺盘孢属真菌有性时期为小丛壳属(Glomerella)[33],在较新近的生物沃斯三域分类系统[34]中棣属于真核域(Eucryota)、真菌界(Fungi)、子囊菌门(Ascomycotina)、盘菌亚门(Arthoniomycetes)、粪壳菌纲(Sordariomycetes)、肉座菌亚纲(Hypocreomycetidae)、小丛壳目(Glomerellales)、小丛壳科(Glomerellaceae),现有的研究发现它们在有性生殖阶段形成子囊壳和子囊孢子[30-33]. 但是,在本研究条件下经过多次培养观察和葡萄园调查,均未见到葡萄炭疽病病原菌的有性时期形态. 因此,今后还需要对此作进一步的观察研究,以期探明葡萄炭疽病菌的有性时期形态,进而弄清葡萄炭疽病的病害循环.
根据需要安全合理地使用高效低毒杀菌剂仍然是当今植物病害综合治理中的重要和必要方法. 对于植物炭疽病,已经有很多关于杀菌剂筛选研究及在作物生产中广泛应用的报道[35-38],市场上也有不少标明专门用于炭疽病控制的药剂种类. 然而,引起炭疽病的刺盘孢属真菌种类很多,一种杀菌剂对不同种类病原真菌引致的炭疽病的药效可能存在较大的差异. 本研究测试了生产中常用的8种杀真菌剂对盘长孢刺盘孢(PT23-01)的抑菌效果,其中25%甲硫乙唑醇SA和30%苯甲吡唑脂SA的抑菌作用很强,对炭疽病的控病率也达到85%和91%,因此可以将它们用于葡萄炭疽病的综合治理措施中,在葡萄生产实践中合理地应用.
综上所述,引起滇西地区葡萄炭疽病的病原菌为3种孢刺盘孢属真菌,其中C. gloeosporiorides是广泛分布并危害的优势种,C. siamense是首次鉴定的葡萄炭疽病的新病原菌;25%甲硫乙唑醇SA和30%苯甲吡唑脂SA是两种高效低毒和无残留的新型复合杀菌剂,对炭疽病菌有很强的抑制作用,所以其对葡萄炭疽病具有非常好的控病效果. 这些研究结果可为该地区葡萄炭疽病的鉴定、诊断、预测预报和绿色综合治理提供重要的科学依据.