


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
随着生活水平的不断提高,人们对猪肉品质的追求也越来越高。猪肉质性状的遗传改良已成为当前的主要研究方向。中国拥有丰富的地方猪资源,这些猪种普遍具有优良的肉质特性,是肉质性状遗传改良的重要遗传素材。深入研究地方猪肉质性状的分子调控机制,挖掘和鉴定与肉质性状相关的调控基因,对于加速肉质性状的遗传改良具有重要意义,也是我国打赢“种业翻身仗”的基础。
猪肉的肌内脂肪(IMF)含量与风味、嫩度和多汁性呈正相关关系,是衡量猪肉品质的关键指标[1-2]。因此,揭示IMF形成和沉积的分子机制对改善猪肉品质至关重要。近年来,关于中国地方猪IMF沉积的遗传机制已有一些研究。例如,对宁乡猪不同发育阶段的肌肉组织进行转录组、miRNA和蛋白组联合分析,发现571对负向互作的miRNA-mRNA以及2对负向互作的miRNA-mRNA-蛋白[3];对调控莱芜猪肌内和皮下脂肪的miRNA和circRNA进行分析,发现circRNA_06 424通过与ssc-miR-339-5p、ssc-miR-744和ssc-miR-328等miRNA竞争性结合,影响PPARD、MMP9、UBA7等靶基因表达,并参与PPAR信号通路、Wnt信号通路、不饱和脂肪酸等信号通路,从而影响莱芜猪IMF和皮下脂肪沉积的差异[4];通过比较淮猪和杜洛克猪不同部位肌肉组织的基因表达差异,分别在背最长肌、腰大肌和肱二头肌中筛选出了175、110和86个差异表达基因[5];利用全基因组关联分析的方法,对453头鲁莱黑猪基因组中与IMF沉积相关的SNP位点进行挖掘,共鉴定出43个IMF沉积相关的SNP位点,注释到42个基因[6];此外,对贵州香猪和大白猪背最长肌之间的转录组差异进行比较分析,鉴定出227个肉质性状相关的候选基因,其中MYOD1、ACTB、ASNS、FOXO1、ARG2、SLC2A4、PLIN2和SCD等29个为贵州香猪肉质性状的关键基因(hub-gene)[7];对快速沉脂阶段(120和240日龄)的皖南花猪背最长肌进行蛋白组学分析,鉴定出106种差异蛋白,并结合转录组数据筛选出7个脂肪沉积候选基因[8];对圩猪和杜洛克猪肌肉组织中的转录组差异进行比较,鉴定出717个差异表达基因,这些差异表达基因与骨骼肌细胞分化、磷脂分解过程和细胞外基质结构成分显著相关[9]。然而,当前对品种内不同个体IMF沉积差异的遗传调控机制还鲜有报道。
盆周山地猪是我国优良地方猪种之一,原产于大巴山、巫山、大娄山、武陵山一带。重庆市主要分布于其辖区的酉阳县(24.6%)、黔江区(22.7%)、彭水县(13.3%)、秀山县(11.4%)、城口县(9.5%)、巫山县(7.5%)及其他相邻区县(11.0%)。四川省主要分布于泸州市(3.4%)、广元市(4.5%)、广安市(4.2%)、达州市(13.4%)、巴中市(21.8%)、宜宾市(17.4%)和南充市(35.3%)等地。2008年起,重庆海林生猪发展有限公司从重庆市酉阳县引入盆周山地猪开展保种和开发利用工作。2014年,依托该公司正式建立盆周山地猪市级保种场,场内采用闭锁繁育方式进行保种,保护群体遗传多样性并维持其遗传结构稳定。盆周山地猪具有沉脂力强、肌纤维细嫩、肉质好、肌肉系水力强等优点[10]。但前期研究发现,盆周山地猪肌肉中IMF含量在群体内的一致性较差,这可能影响其在优质猪肉生产中的进一步应用。因此,揭示盆周山地猪IMF沉积差异的分子调控机制对其肉质改良至关重要。然而,目前尚未有针对盆周山地猪IMF沉积差异分子机制的研究。本研究以高、低IMF含量的盆周山地猪为研究对象,利用高通量测序技术对两者背最长肌进行转录组测序分析,筛选两者差异表达的mRNA和lncRNA,并对这些差异mRNA和lncRNA进行生物信息学分析,筛选出影响盆周山地猪IMF含量的关键候选mRNA和lncRNA。研究结果加深了对盆周山地猪IMF沉积的理解,也为盆周山地猪肌肉品质的选育提供了分子依据。
全文HTML
-
实验用相同日龄的26头盆周山地猪个体选自盆周山地猪市级保种场,并在保种场内统一条件下饲养、自由采食和饮水。达90±5 kg体重时在重庆市涪陵区南沱镇屠宰场屠宰,统一采集2~3腰椎处的背最长肌,用于后续IMF含量测定、RNA提取、油红O染色。其中,RNA提取样本迅速置于液氮中,运回实验室后于-80 ℃冰箱保存;油红O染色样本用4%多聚甲醛固定;剩余样本于低温环境运送至农业农村部种猪质量监督检验测试中心(重庆),打碎、称重后-20 ℃保存,用于IMF测定。
-
根据中华人民共和国农业行业标准《猪肉品质测定技术规程》 (NY/T 821—2019)的要求,采用索氏浸提法进行IMF含量的测定。依据每个样品IMF含量的测定值,2.5%为极低IMF含量组的筛选阈值,5.0%为极高IMF含量组的筛选阈值筛选出盆周山地猪群体内极低IMF含量组个体3头、极高IMF含量组个体5头。
-
通过油红O对目标组织进行染色,可以确定目标组织中脂类物质沉积的情况。具体步骤如下:将采集的背最长肌修剪成2 mm左右的长度后放入OCT包埋胶中,于恒温冰冻切片机中进行包埋。待OCT胶凝固后,于恒温冰冻切片机中连续切片,制成厚度为6 μm的切片。将切片于甲醛中固定10 min后用蒸馏水清洗,再用60%的异丙醇浸洗。随后,将切片浸泡在油红O染液中10 min后,再将切片浸入60%异丙醇分色至背景无色。然后,用蒸馏水清洗切片后用苏木精复染1 min。最后,用蒸馏水清洗切片,待切片晾干后用甘油封片。
-
利用Trizol试剂提取每个背最长肌样本的总RNA,用Nanodrop 2000检测RNA的浓度和纯度,1%琼脂糖凝胶电泳检测RNA的完整性,Agilent 2100检测样本的RIN值。对检测合格的样品利用Illumina PE150测序平台进行建库和测序,相关工作在北京百迈客生物科技有限公司完成。
-
利用FastQC软件,采用如下质控标准对原始数据进行质量控制:①去除含接头的reads;②去除质量低的reads(质量值Q≤20的碱基数占整条reads的50%以上);③去除片段长度小于50 bp的reads;④去除含N比例大于10%的reads。利用STAR软件将质控后的高质量数据比对到猪的参考基因组(Sscrofa 11.1)上。利用Cufflinks(2.2.1)软件对比对到基因组的数据进行组装。随后,用Assemblyline软件对转录本进行单外显子的过滤,并通过TACO对所有转录本进行合并,筛选去除含有clipped exon(exon长度小于15个碱基)、FPKM(每百万映射读取的每千碱基片段数,Fragments Per Kilobase of transcript per Million mapped reads)低于0.1以及长度小于250个碱基的转录本。再通过与基因组对比,判断是否有已注释基因或转录本的存在。对于不能被基因组注释的转录本,通过CPC2软件判断其蛋白编码潜能(编码或非编码)。接着,用Pfamscan软件分别对所有编码和非编码的转录本进行蛋白数据库(Pfam31)的比对,分别统计每个蛋白域比对上的转录本(编码或非编码)数量,并通过fisher方法检验编码与非编码转录本都比对上的蛋白域的相似性(当p>0.05或OR<10时,认为非编码对应的蛋白域更趋向于编码,其中OR为优势比),没有被基因组注释的编码转录本即为TUCP(Transcripts with Unknown Coding Potential),其余为lncRNA。最后通过kallisto软件对mRNA和lncRNA进行定量和count数目的统计。
-
利用edgeR软件包进行mRNA及lncRNA的差异表达分析,以|log2FC|>0.75、FDR<0.05为标准筛选差异表达基因,其中FC(Fold Change)为差异倍数,FDR为(False Discovery Rate)阳性错误率。利用Metascape网站(
https://metascape.org/gp/index.html#/main/step1 )对差异表达基因进行GO(Gene Ontology,基因本体论)和KEGG(Kyoto Encyclopedia of Genes and Genomes,京都基因与基因组百科全书)功能富集分析,以p<0.01为显著富集条目和通路。 -
将lncRNA基因组位置上下游100 kB的基因认为是lncRNA的顺式作用靶基因。反式作用靶基因预测的基本原理认为lncRNA的功能跟编码基因的位置没有关系,而与其共表达的蛋白编码基因相关。可以通过样本间lncRNA与蛋白编码基因的表达量相关性分析来预测其靶基因。将lncRNA-mRNA之间的Pearson相关系数大于0.9或小于-0.9但p<0.05的基因判定为lncRNA的反式作用靶基因。
-
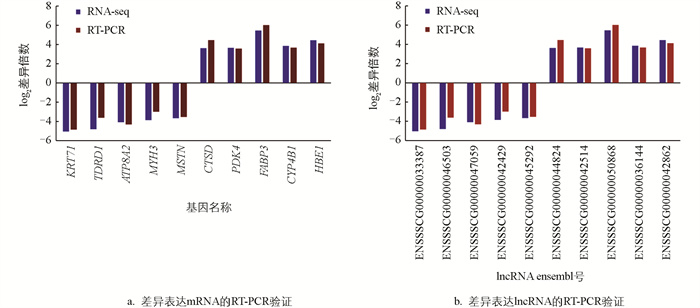
为了验证测序结果的可靠性,分别挑选10个差异表达的mRNA和lncRNA(上调、下调各5个)进行RT-PCR验证。使用β-actin基因作为内参基因,所有引物信息见表 1,所有引物均在苏州金唯智生物科技有限公司合成。利用2-ΔΔCt方法计算基因的表达量。
1.1. 实验材料
1.2. IMF含量测定
1.3. 油红O染色
1.4. 总RNA提取及建库
1.5. 数据的质控、比对及组装
1.6. mRNA及lncRNA的差异表达分析
1.7. 差异表达lncRNA的靶基因的预测
1.8. 测序结果可靠性分析
-
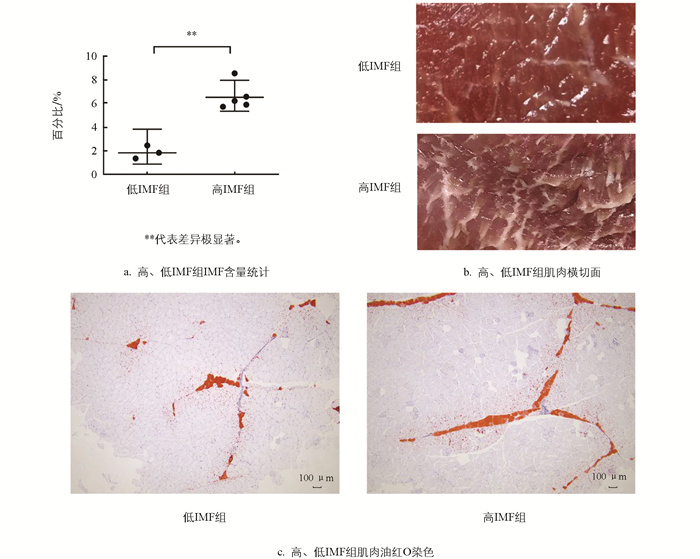
利用索氏抽提法进行IMF测定,依据IMF脂肪含量测定结果,分别命名为高IMF组和低IMF组,其中高IMF组平均IMF含量为6.58±1.14%,低IMF组平均IMF含量为1.87±0.55%(图 1a)。高、低IMF组肌肉横切面及油红O染色结果如图 1b、1c所示,表明高IMF组肌肉组织内IMF分布比低IMF组更广。
-
对测序结果进行质控,共获得8个样本总计157.78 GB的clean reads,每个样本至少获得17.48 GB的clean reads。将clean reads比对到猪参考基因组(Scrofa 11.1)发现,样本的总比对序列数占比为96.82%~97.83%,唯一比对序列数占比为78.92%~89.91%,多重比对序列数占比为7.55%~17.90%(表 2),测序数据质量整体较好,可以进行后续分析。
-
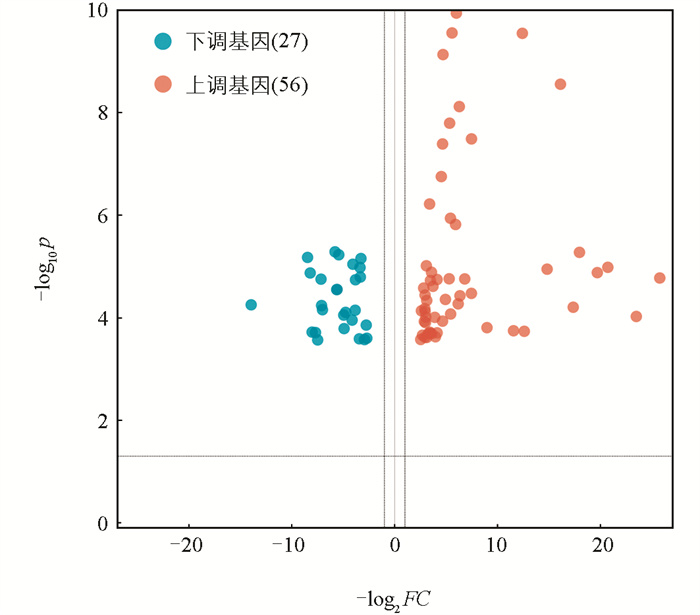
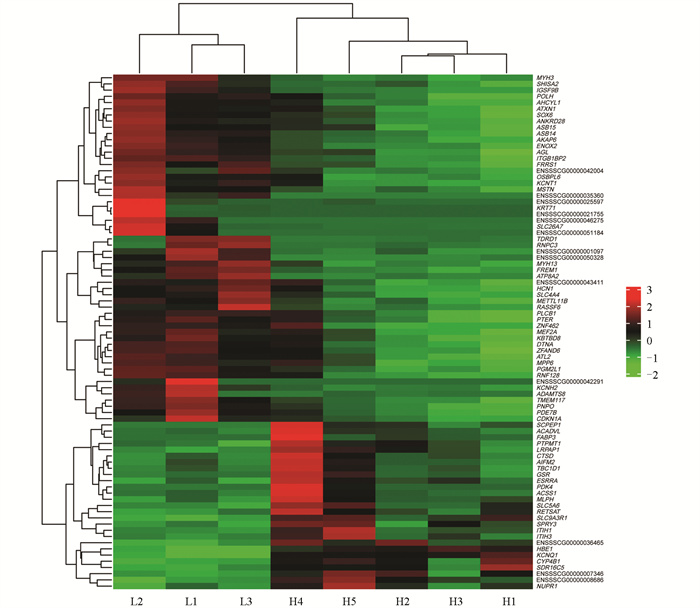
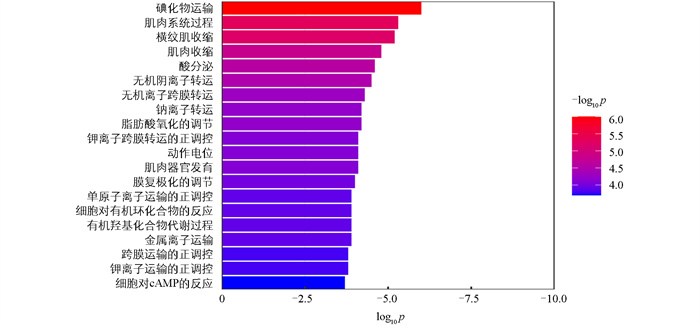
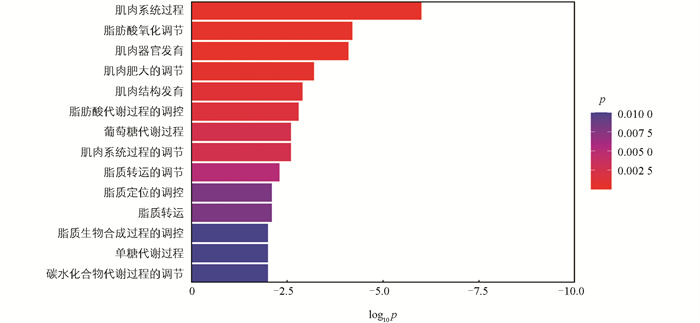
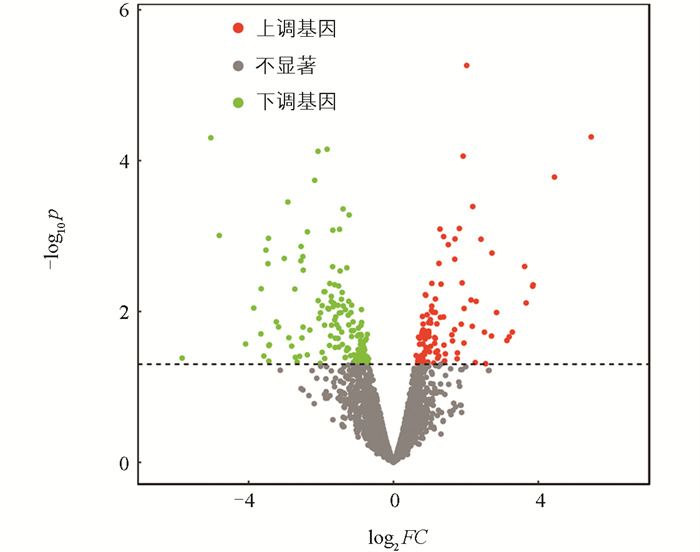
利用edgeR软件包进行mRNA筛选,以|log2FC|>0.75、FDR<0.05为显著表达差异条件。结果表明高、低IMF组之间共有83个差异显著mRNA,相对于高IMF组,低IMF组中有56个mRNA显著上调,27个mRNA显著下调(图 2),主要涉及的相关基因有MYH3、MYH13、PDK4、MSTN、FABP3等(图 3)。对差异表达基因进行GO条目分析发现,共有111个条目显著富集(p<0.05),主要参与肌肉系统过程、脂肪酸氧化调控、肌肉器官发育等生物学过程(图 4)。这些显著富集的GO条目中,14个与肌肉和脂肪相关,主要参与肌肉发育、脂肪酸氧化、脂肪酸代谢、葡萄糖代谢、脂质转运等过程(图 5)。KEGG功能富集分析结果表明,这些差异基因显著富集的信号通路有4个,即:甲状旁腺激素的合成、分泌和作用,胃酸分泌,糖尿病性心肌病以及胰液的分泌。
-
以|log2FC|>0.75、p<0.05为显著表达差异条件筛选差异lncRNA。结果表明,两组之间共有237个已知的差异lncRNA。相较于高IMF组,低IMF组中有144个lncRNA显著下调,93个lncRNA显著上调(图 6)。
-
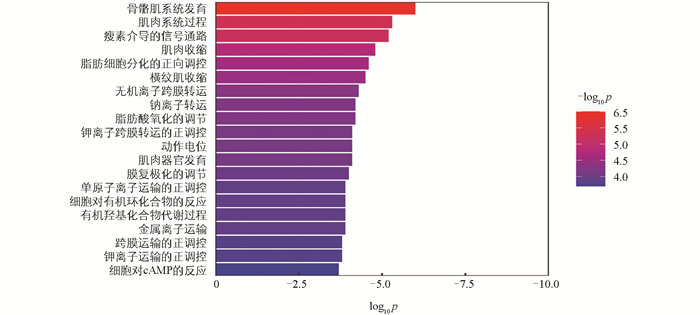
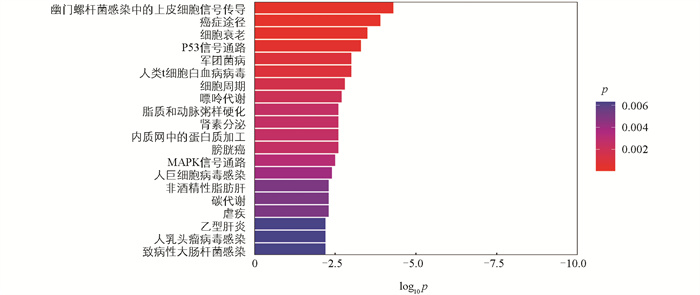
基于lncRNA在染色体上的位置,在差异显著lncRNA的上下游100 kB范围内共找到KIF5A、ACOX3、PDK4、KLF12、KLF3、LEP等413个编码基因(顺式作用靶基因)。GO富集分析发现,这些基因在405个GO条目中显著富集,包括骨骼肌系统发育、瘦素介导的信号通路、脂肪细胞分化的正向调控等生物学过程(图 7)。KEGG信号通路分析结果表明,这些基因富集在细胞周期、细胞衰老、脂质和动脉粥样硬化及非酒精性脂肪肝等信号通路(图 8)。基于lncRNA-mRNA之间的Pearson相关系数,在这些差异显著的lncRNA中,共鉴定得到11个lncRNA与515个编码基因存在相关关系。其中有6个差异显著的lncRNA与11个差异显著的编码基因之间存在相关关系。此外,有2个差异显著的lncRNA与3个差异显著的肉质相关编码基因存在潜在的靶向调控关系(表 3)。
-
为了验证转录组测序结果的准确性,随机选取10个mRNA和lncRNA进行实时荧光定量PCR验证。结果发现,所验证的mRNA和lncRNA(低IMF组vs高IMF组)表达趋势与转录组测序的表达趋势(低IMF组vs高IMF组)一致(图 9),表明此次转录组测序结果可靠性高,可以用于后续研究。
2.1. 盆周山地猪IMF含量测定及形态学分析
2.2. 测序结果的比对分析
2.3. 基因差异表达及功能分析
2.4. 差异lncRNA表达分析
2.5. lncRNA的靶基因预测
2.6. 测序结果的实时荧光定量PCR验证
-
盆周山地猪是我国优良地方猪种之一,平均IMF含量与肌肉的理想IMF含量3%~5%相符,是优良的育种素材,但是其IMF含量在群体内的一致性较差,这将对后期的利用产生影响。本研究以这一问题为出发点,通过比较高、低IMF含量猪只背最长肌的mRNA和lncRNA表达差异,筛选盆周山地猪IMF沉积差异的候选mRNA和lncRNA,以期为盆周山地猪肉质的选育提供分子依据。本研究首先通过索氏抽提法结合油红O染色在盆周山地猪群体中筛选出了高IMF组和低IMF组,高IMF组平均IMF含量(6.58±1.14%)是低IMF组(1.87±0.55%)的3.5倍(图 1a),说明在盆周山地猪群体中个体之间IMF的含量有很大差异,有必要进一步挖掘调控盆周山地猪IMF沉积的关键基因。
随后对高IMF组和低IMF组肌肉组织进行了转录组测序,测序结果发现高IMF组和低IMF组之间共有83个差异表达的mRNA,推测这些差异表达基因可能与两组猪IMF含量差异相关,为了探究这些基因的生物学功能,对这些基因进行了GO功能富集分析,分析结果中值得注意的有脂肪酸氧化调节、脂肪酸代谢过程、葡萄糖代谢过程、脂质转运、脂质生物合成过程、单糖代谢过程、碳水化合物代谢过程等与能量代谢相关的过程,推测高IMF组和低IMF组猪IMF沉积能力的差异是由脂质、能量代谢相关基因的差异表达造成的。转录组测序结果还发现高IMF组和低IMF组之间有237个差异表达的lncRNA,为了明确差异表达lncRNA的潜在功能,对这些差异表达lncRNA进行了顺式作用靶基因的预测,共预测出413个基因,并对这些靶基因进行了GO功能和KEGG通路富集分析。GO分析结果显示这些靶基因主要富集在骨骼肌系统发育、瘦素介导的信号通路、脂肪细胞分化的正向调控等生物学过程。KEGG发现这些靶基因主要富集在细胞周期、细胞衰老、脂质和动脉粥样化及非酒精性脂肪肝等信号通路。基于lncRNA-mRNA之间的Pearson相关系数,筛选出有2个差异显著的lncRNA与3个差异显著的肉品质相关编码基因存在潜在的靶向调控关系,这3个基因分别为FABP3、PDK4和CTSD。因此推测,lncRNA通过调控这些基因的表达影响盆周山地猪IMF的沉积过程。
腺苷三磷酸酶家族中有4种丙酮酸脱氢酶激酶(Pyruvate Dehydrogenase Kinase,PDKs)同工酶,分别为PDK1、PDK2、PDK3和PDK4 [11]。PDK4参与动物体内三羧酸循环、脂肪代谢、糖酵解、ATP的形成等多个新陈代谢过程[12],在人和小鼠的心脏、肝脏、骨骼肌和脂肪组织中有较高表达量[11]。在家畜各组织中,PDK4基因在肌肉和脂肪组织中表达量较高,在屯昌猪背最长肌中表达量最高[13],在陆川猪和大白猪的皮下脂肪表达量最高[14-15],在巴马香猪和杜长大猪的腹脂中高表达[16],提示PDK4在家畜肌肉和脂肪生长过程中具有重要作用。已有研究发现,PDK4的表达量与鸡的脂肪沉积速度相关[17],PDK4的过表达能够增加猪的脂肪代谢效率[15],且PDK4基因的SNP位点与IMF、肌肉系水力等肉质性状具有相关性[18-19]。PDK4可以抑制丙酮酸向乙酰CoA转化[20],乙酰CoA减少后会引起脂肪酸氧化抑制剂丙二酰辅酶A的减少,说明PDK4的上调表达可以促进脂肪酸氧化[21]。PDK4敲基因小鼠禁食后,随着血糖降低,脂肪酸氧化速率降低,葡萄糖和丙酮酸氧化速率增加,且增加速率一致[22]。结合以上的研究结果和本研究的结果,推测PDK4基因可能是影响盆周山地猪IMF沉积的关键候选基因。
脂肪酸结合蛋白(Fatty Acid Binding Protein,FABPs)作为脂质结合蛋白超家族的成员,广泛参与动物体脂质代谢和脂肪沉积等过程[23],对脂肪酸具有很强的亲和性[24],其中FABP3基因目前广泛被认为是调控IMF沉积的候选基因,主要通过参与细胞中长链脂肪酸的摄取、转运和利用实现脂肪代谢调节[25],目前有大量研究证实FABP3基因多态性与猪IMF之间存在显著相关性[26-28]。本研究的转录组测序结果发现,盆周山地猪高IMF组和低IMF组的背最长肌FABP3基因mRNA的表达差异显著,推测FABP3基因是调控盆周山地猪IMF沉积的重要基因。
-
本研究通过转录组测序在盆周山地猪高IMF组和低IMF组的背最长肌中共鉴定出了83个差异表达mRNA和237个差异表达的lncRNA。差异表达lncRNA和mRNA主要在脂质、能量代谢相关的信号通路中富集。lncRNA-mRNA互作分析筛选出了2个差异显著的lncRNA(ENSSSCG00000044255、ENSSSCG00000048256)与3个差异显著的肉质相关编码基因(FABP3、PDK4和CTSD)存在潜在的靶向调控关系,因此推测这些lncRNA和mRNA是影响盆周山地猪IMF沉积的重要候选基因。