


 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
由于农业生产中含氮化肥的过量施用、生活污水的大量排放及工业废水的直接排放等因素,水体硝酸盐污染日益严峻。硝酸盐在水体中的累积不仅会引发富营养化,破坏水生生态系统结构与功能,其不完全还原产物(亚硝酸盐)还会诱发高铁血红蛋白血症,对人类健康构成极大威胁。硝酸盐作为一种氧化态电子受体,其去除需依赖电子供体的还原。一些低C/N废水(一般指C/N<8)[1],例如生活污水、市政污水处理厂二沉池出水等,因有机碳源匮乏,导致硝酸盐无法被完全去除。为高效去除该类废水中的硝酸盐,传统异养反硝化工艺常依赖大量外源有机碳源的投加,这不仅大幅提升了处理成本,又易因碳源利用不充分造成二次污染。因此,开发高效、经济、环境友好的低C/N废水脱氮技术,已成为水环境修复领域的核心需求[2-3]。
微生物电化学系统(Microbial Electrochemical System,MES)作为融合微生物代谢与电化学过程的绿色创新技术,为低C/N废水脱氮提供了极具前景的解决方案。与传统工艺不同,MES以电极为电子供体驱动反硝化菌生长,大幅降低了反硝化过程对有机碳源的依赖,契合绿色低碳治理理念[4-5]。然而,该系统仍面临电极与生物膜间胞外电子传递效率偏低的难题,反硝化菌难以高效获取电子,限制了硝酸盐还原速率[4]。在MES阴极反硝化体系中,电活性生物膜是电子传递与硝酸盐还原的核心载体。已有研究证实,通过电极极性反转方式可获得双向电子转移生物膜,其中的电活性菌可通过胞外电子传递搭建电极与反硝化菌间的电子转移桥梁,并驱动硝酸盐逐步还原为氮气。然而,电极极性的切换易引起活性细菌的大量流失,导致系统脱氮效能难以维持,影响长期运行稳定性[6]。
优化电极表面特性及系统运行策略可实现对电活性生物膜的有效调控。利用功能性纳米材料修饰电极,可精准调控其电化学特性与表面微观结构:一方面可提升电极导电性、电容性能与电子储存能力,强化电极与生物膜间的电子传递效率[7];另一方面可为电活性细菌提供适宜附着位点,促进功能菌群富集,优化微生物群落组成与生物膜空间结构[8-9]。周期极性反转策略通过交替切换电极的阴阳极属性,定向培养双向电子传递生物膜,使其在阳极期储存电子、阴极期释放电子还原硝酸盐,实现电子的高效利用[4, 6]。二氧化锰(MnO2)作为典型的过渡金属氧化物,兼具优异的导电性、电容特性、化学稳定性与生物相容性,已被广泛应用于电催化与生物电化学领域[9]。已有研究表明,MnO2可刺激电活性菌产生胞外囊泡[10-11],增强电极与电活性菌间的电子转移。同时,其赝电容特性也有助于增强种间电子转移能力,提升厌氧系统的产甲烷效率[12]。因此,将MnO2修饰于碳布电极表面,既可提升电极电子储存与传递能力,又能优化电极表面形貌,为Geobacter sp.等电活性细菌的定殖营造适宜微环境,促进电活性生物膜的形成与功能发挥,提高系统稳定性。
本研究通过水热合成法制备MnO2修饰碳布电极,构建单室MES反应器,采用周期极性反转策略定向培养双向电子传递生物膜,以未修饰空白碳布电极为对照,系统探究MnO2修饰对MES处理低C/N废水反硝化效能的影响。从脱氮效能、电子传递特性、微生物群落结构及生物膜形貌组成等角度,解析MnO2修饰的强化机制,揭示其对电极-生物膜电子传递、生物膜结构优化及功能菌群富集的调控规律,旨在为提升MES脱氮效能提供可行技术手段,并为功能性电极材料开发与运行策略优化提供理论依据和实验支撑。
全文HTML
-
通过水热合成法制备MnO2修饰碳布工作电极,具体流程如下:在磁力搅拌条件下,将256 mg氟化钾(KF)和200 mg高锰酸钾(KMnO4)依次加入20 mL去离子水中,搅拌溶解后加入25 mL 0.5mol/L的H2SO4溶液,继续搅拌30 min。随后,取一块碳布放入上述溶液中,将溶液和碳布同时移入内衬聚四氟乙烯的不锈钢高压反应釜中,于120 ℃反应80 min。待反应完成后,将反应釜冷却至室温,取出碳布,用去离子水洗涤3次,而后于60 ℃真空干燥箱中干燥12 h,即得到MnO2修饰的工作电极。
反应器的组装:使用有机玻璃构建立方体单室生物反应器,其内部空腔横截面直径约为35 mm,腔体长度为30 mm,反应器有效容积为28 mL。采用合成的MnO2修饰碳布(直径30 mm)作为工作电极,安装相同直径的钛网(120目,纯度99.99%,昆山恒博新材料有限公司)作为对电极。参比电极为饱和甘汞电极(SCE,相对标准氢电极的电位为+241.5 mV),固定在工作电极旁。
反应器的运行:启动阶段,反应器进水中加入体积分数为20%的生活污水,并在+0.2 V vs SCE电位下运行。在阳极电流输出稳定后,为培养双向电子传递生物膜,对反应器施加-0.5 V vs SCE的恒定负电位。阳极期以乙酸钠(1 g/L)为电子供体,电极为电子受体。阴极期进水模拟二沉池出水,提供2 mmol/L的硝酸盐氮作为电子受体,电极为唯一电子供体。运行过程中,通过恒电位仪(CHI 1000C,上海辰华仪器有限公司)施加恒定电位。此外,阳极和阴极电解质溶液均含有50 mmol/L PBS缓冲液、1.25%微量矿物质溶液和0.50%维生素溶液[13]。进水前,阳极和阴极进水均用N2(99.99%)曝气30 min以保持厌氧状态。阳极、阴极期运行时间均为1 d,待获得成熟阳极生物膜后,系统共运行了14个完整的阴阳极周期。
-
化学需氧量(Chemical Oxygen Demand,COD)的测定:COD采用《水质化学需氧量的测定快速消解分光光度法》(HJ/T399—2007)测定。使用移液枪准确量取2 mL待测溶液置于COD测试管中,随后准确量取1 mL消解液1,之后准确加入4 mL消解液2,拧紧瓶盖,充分震荡使管内液体混合均匀。在快速消解仪DRB 200上设定恒温消解程序,消解温度为165 ℃。待仪器升温完成后,将消解管置于消解槽中恒温消解15 min,之后缓慢降温,待温度降到120 ℃时将消解管取出,震荡混合均匀,置于室温下自然冷却至常温。随后用分光光度计DR 3900测定其吸光度。
氮含量的测定:测定前,水样先经过0.45 μm滤膜过滤。硝酸盐氮(NO3--N)使用紫外分光光度法测定;亚硝酸盐氮(NO2--N)使用N-(1-萘基)-乙二胺光度法测定;氨氮(NH4+-N)使用纳氏试剂分光光度法测定,由于系统中游离氨浓度仅占总氨氮的0.8%~1.0%,氨氮基本以NH4+-N形式存在,为简化表达,使用NH4+-N来代替氨氮。总氮去除率通过式(1)计算:
微生物群落结构分析:于第14个运行周期结束后采集两组电极表面生物膜样品,液氮速冻后置于-80 ℃冰箱保存。对生物膜样品进行16S rRNA高通量测序,探究其微生物群落结构组成。具体流程如下:首先对样品进行DNA提取,测试DNA浓度后,利用1%琼脂糖凝胶电泳检测DNA的质量,若电泳条带清晰明亮即可进行下一步扩增。对细菌16S rRNA基因的V3-V4区进行扩增,正向引物为338F(5′-ACTCCTACGGGAGGCAGCAG-3′),反向引物为806R(5′-GGACTAACHVGGGTWTCTAAT-3′)[14],经变性、退火、延伸后得到PCR产物。将PCR产物回收并进行纯化处理后,进行DNA浓度检测及定量。依据Illumina Miseq平台的操作规范,对纯化后的DNA片段构建基因文库。高通量测序完成后,利用美吉生物公司云平台(
www.i-sanger.com )进行生物信息学分析,以优化后的扩增子序列变体特征表为基础,在属分类水平下,对空白碳布组和MnO2修饰碳布组样品进行群落结构可视化分析。同时,基于FAPROTAX方法,在该云平台完成微生物群落的氮代谢功能预测。电化学测试方法:进行有底物状态下的循环伏安(Cyclic Voltammetry,CV)扫描时,待一个运行周期结束后,向反应器中加入新鲜的电极液(与进水阳极液保持一致),并在开路状态下稳定2h,使用电化学工作站的CV模块进行测试。通过对CV曲线各数据点电流(I)对应的电极电位(E)求一阶导数(DCV),进而获得有底物状态下电活性生物膜的催化活性位点。在进行无底物状态下的CV扫描时,待各系统完成一个完整周期运行并消耗完底物后,加入50 mmol/L PBS缓冲液,开路稳定2 h以上再进行测试。CV扫描的参数设置如下:起始电位-0.5 V;终止电位+0.6 V;有底物CV测试扫描速率为5.0 mV/s,无底物CV测试扫描速率分别为0.5、0.7和1.2 mV/s;扫描圈数为3圈,选择最后一圈结果作为最终数据。
生物膜三维结构分析:使用激光共聚焦显微镜(CLSM 800,德国蔡司)对生物膜三维结构进行观察,观察前对生物膜进行活死菌染色。染色母液配制:将1 mg钙黄绿素AM溶于1 mL二甲基亚砜中,1 mg碘化丙啶溶于1 mL去离子水中。工作液配置前,先将50 mmol/L PBS缓冲液置于37 ℃恒温箱中备用,再分别配制钙黄绿素AM工作液(取2 μL母液稀释于1 mL PBS缓冲液中)与碘化丙啶工作液(取1 μL母液稀释于1 mL PBS缓冲液中)。随后用恒温PBS缓冲液冲洗待测电极生物膜,除去电极表面多余的培养基,先加入钙黄绿素AM工作液,37 ℃染色15 min,再加入碘化丙啶工作液,37 ℃继续染色5 min,之后用恒温的PBS缓冲液快速冲洗电极生物膜2次,再浸泡电极5 min以去除多余染液。染色完成后,将电极生物膜置于激光共聚焦显微镜下进行三维形貌测试。生物膜胞外聚合物(Extracellular Polymeric Substance,EPS)分析:首先对生物膜EPS进行染色,荧光染料异硫氰酸荧光素(Fluorescein Isothiocyanate,FITC)用于蛋白染色,染料刀豆蛋白A(Concanavalin A,ConA)和钙荧光白(Calcofluor White,CFW)分别用于测试α-D-吡喃葡萄糖和β-D-吡喃葡萄糖。染料工作液的配置参考Liang等[15]的方法,全程在暗室进行,具体染色步骤如下:首先进行样品固定,将附着有电活性生物膜的电极浸泡在2.5%戊二醛溶液中,于4 ℃下固定12 h,随后倒掉多余戊二醛溶液,向电极生物膜样品中加入0.1 mol/L NaHCO3溶液(pH=9.0)至完全覆盖样品,室温反应10 min后倒掉溶液;然后进行样品染色,将FITC溶液加入样品并覆盖电极生物膜,室温反应1 h,用PBS缓冲液缓慢冲洗后,加入ConA溶液染色30 min,再用PBS缓冲液轻微冲洗电极,随后加入CFW,室温反应30 min;最后用PBS缓冲液缓慢冲洗掉多余染色剂。测试时,将样品置于激光共聚焦显微镜下观测,通过设置Z轴层扫对EPS形态进行三维建模。
-
电流积分:阳极期,电极作为电子受体,产生正电流;阴极期,电极作为电子供体向生物膜提供电子,产生负电流。电流积分结果可反映电极与生物膜之间的双向电子传递能力,计算公式如下:
式中:nA-anode为阳极期电极获得的电量(mmol);nA-cathode为阴极期电极提供的电量(mmol);IAn和ICath分别为阳极期和阴极期记录的正电流和负电流(A);t为采集一个电流数据的时间间隔(s);F为法拉第常数(96 485 C/mol)。
电子平衡分析:在阴极期,电极是唯一的电子供体,在阳极期,电子从乙酸盐氧化中释放。其中一部分电子通过电流形式转移至电极,另一部分则储存于生物膜中,用于阴极期的硝酸盐还原过程[16]。因此,在阴极期,总电子量来自电极和生物膜,计算公式如下:
式中:nC为阴极期总电子量(mmol);nA-cathode为阴极期电极提供的电子量(mmol);nbiofilm为生物膜储存的电子量(mmol)。
各系统中用于硝酸盐还原的电子量计算公式如下:
式中:nN为用于硝酸盐还原的电子量(mmol);ΔNO3--N为去除的硝酸盐氮摩尔数;ΔNO2--N为每个循环后出水中产生的亚硝酸盐氮摩尔数;ΔNH4+-N为出水中产生的氨氮摩尔数。
直接电子传递(Direct Electron Transfer,DET)和间接电子传递(Mediated Electron Transfer,MET) 对电流构成的贡献:为明确电极与生物膜间主导的电子传递过程,对不同扫描速率下无底物循环伏安(CV)测试所得数据进行线性拟合分析,计算公式如下:
式中:i为特定外加电位下的峰值电流(包括阳极峰值电流和阴极峰值电流,mA);v为扫描速率(mV/s);b为斜率,是评估主导电子传递过程的重要指标;k为常数。当b≈1时,对应直接电子传递过程的动力学控制反应;当b=0.5时,表现为扩散控制行为,对应介导电子传递过程;当0.5<b<1时,代表电子传递过程为动力学控制和扩散控制相结合[17]。
在特定扫描速率下进一步分析了DET(动力学控制过程)和MET(扩散控制过程)的贡献。电流i由与扫描速率(v)和扫描速率平方根(v1/2)相关的两部分组成,如式(7)所示:
式中:k1和k2是常数;k1v代表动力学控制过程的贡献;k2v1/2表示扩散控制过程的贡献。将式(7)两边除以v1/2,得到式(8):
通过在不同扫描速率下对i/v1/2和v1/2进行线性拟合,可以得到斜率k1。
DET贡献率计算公式如下:
为了确定MnO2修饰组与空白对照组在总氮去除率上是否具有统计学意义,用Microsoft Excel 2019软件进行了t检验,当p<0.001时,判定两组数据间差异有统计学意义。
1.1. 反应器的组装与运行
1.2. 分析方法
1.3. 计算
-
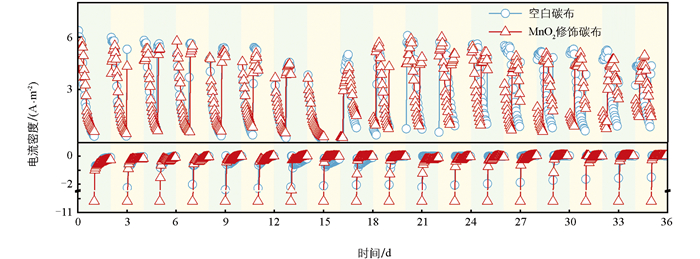
采用周期性极性反转方法,成功获得了具有双向电子传递功能的生物膜。结果表明,阳极期和阴极阶段分别产生正向和负向电流,说明电活性生物膜与电极之间实现了双向电子传递。如图 1所示,系统在经历多次极性反转后,阳极最大输出电流密度及阴极输出电流密度在各周期间并未产生巨大变化,说明该生物膜的双向电子转移能力较为稳定。系统阳极期的最大电流密度均可维持在约5.9 A/m2,MnO2修饰组在阴极期的起始电流密度较空白碳布组更低,约为-10.2 A/m2。电极极性反转后,生物膜内储存的电子可能发生瞬时释放,且电极表面生物膜电容强度越大时,其产生的初始电流绝对值越高。在本系统中,MnO2修饰碳布组的初始电流值显著高于对照组,这与其生物膜电量存储能力直接相关,得益于MnO2的电容特性,使得生物膜具备暂时储存更多电子的能力。
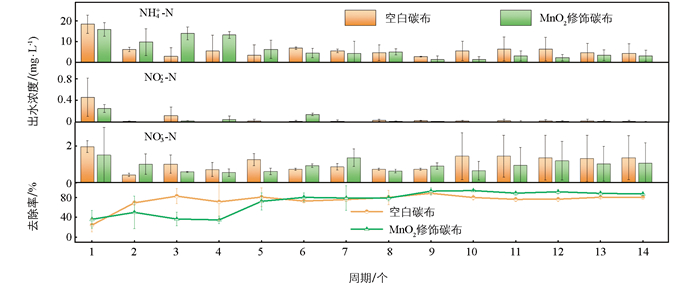
对极性反转后生物膜的脱氮能力进行探究,图 2为MnO2修饰组和对照组在阴极状态运行24 h后的硝酸盐转化情况。结果显示,两组均未观察到明显的亚硝酸盐积累;NH4+-N均呈先累积后降低的趋势,这可能与阳极期富集的高丰度Geobacter sp.有关,与部分生物电化学系统中存在的硝酸盐还原为氨氮的现象类似[18]。在硝酸盐的持续胁迫下,系统反硝化功能逐步被驯化,生物膜的代谢路径发生功能性调整。最终,MnO2修饰组总氮去除率由初始的36%提升到了91%,显著高于空白碳布组(p<0.001),表明MnO2修饰对生物膜功能演替起到正向调节作用,有利于微生物群落的稳定发展。
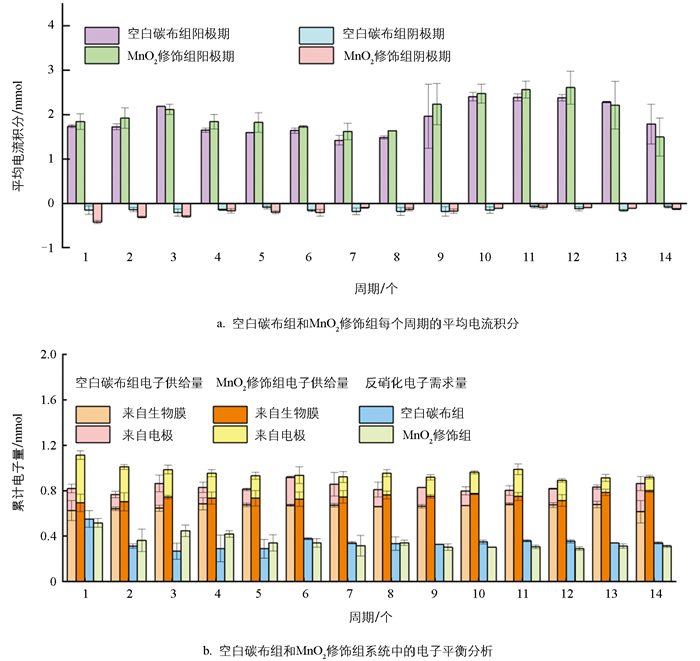
已有研究证实,电活性生物膜具有“生物赝电容”特征,能够在阳极期将电子储存在聚羟基脂肪酸酯(Polyhydroxyalkanoate,PHA)、胞外聚合物(EPS)或细胞色素C中,并在电子供体缺乏的阴极期按需释放[19]。图 3a展示了空白碳布组与MnO2修饰组在14个运行周期内阳极期与阴极期的平均电流积分,直观呈现了系统电子储存与释放的动态特征。空白碳布组与MnO2修饰组阳极期电流积分均显著高于阴极期,证实阳极期是系统电子储存的关键时期;MnO2修饰组阳极期平均电流积分在多数周期内高于空白碳布组,充分体现了MnO2的电容特性。MnO2作为典型赝电容材料,可通过Mn4+/Mn3+的可逆氧化还原反应直接储存电子。同时,MnO2修饰组阴极期电流积分的绝对值也略高于空白组,表明其在阴极期可更高效地释放储存电子,为硝酸盐还原提供电子供体。空白碳布组和MnO2修饰组生物膜在阳极期的最大电子储存量分别约为(0.69±0.01) mmol和(0.80±0.03) mmol,这部分电子可作为阴极期硝酸盐还原的补充电子供体。图 3b的电子平衡分析进一步显示,在所有运行周期中,MnO2修饰组来自生物膜与电极的累计电子量均显著高于空白碳布组,这是其电子储存能力提升的直接量化体现。其中,生物膜贡献电子量的增加,可能与MnO2诱导电活性菌(如Geobacter属)过表达C型细胞色素等关键蛋白有关,增强了生物膜的电子储存与传递效能;电极贡献电子量的提升,则应归因于MnO2材料自身赝电容特性所贡献的额外电子储存,弥补了空白碳布电极因缺乏电容特性而导致储电能力不足的局限。与此同时,MnO2修饰组用于反硝化的电子量持续高于空白碳布组,这有利于硝酸盐的还原。周期性极性反转策略利用“生物赝电容”特性,实现了充电和放电反应的交替进行,有利于双向电子传递生物膜的形成。在本系统中,硝酸盐还原的理论电子需求量略小于电极供给电子与生物膜储存电子之和,推测多余电子可能用于生物同化作用。
-
MnO2修饰碳布电极对微生物群落结构与氮代谢功能基因的表达有显著调控作用。对系统运行结束后的群落结构分析表明,MnO2修饰实现了对功能菌的选择性富集与筛选,核心电活性菌Geobacter sp.作为电子传递关键功能菌[20],其相对丰度从空白对照组的50.84%升至MnO2修饰组的70.21%(图 4a),该菌可通过外膜细胞色素C与导电菌毛构建电极与微生物间的直接电子传递路径[21],为反硝化过程持续提供电子供体。另一电活性菌Geothrix sp.的相对丰度从空白对照组的1.16%升至MnO2修饰组的1.45%,进一步完善了生物膜胞外电子传递网络[22];Thiobacillus sp.是典型的电极营养型自养反硝化菌,在活性炭、磁铁矿等的作用下可与Geobacter sp.形成互营关系,实现种间电子传递[23]。反硝化菌Thauera sp.可高效将硝酸盐还原为氮气[24],其相对丰度在修饰组中保持稳定,保障了反硝化功能的稳定性;修饰组中反硝化菌Pseudazoarcus sp.丰度虽低但略有提升(从0.02%提升至0.04%),该菌可高效利用电极型无机电子供体进行反硝化[25]。与此同时,MnO2修饰组中Dechlorobacter sp.等反硝化菌相对丰度下降,Pseudomonas sp.和Thiopseudomonas sp.未检出,说明MnO2修饰降低了菌群整体的多样性,提升了群落功能专一性。此外,未分类杂菌丰度从空白对照组的29.74% 降至MnO2修饰组的23.17%,进一步增强了群落功能专一性。
已有研究表明,Geobacter sp.作为“电子桥梁”,可通过胞外电子传递系统将电极表面电子传递至Thauera sp.等核心反硝化菌,构建起电活性菌、反硝化菌互惠协同的功能体系[26]。与空白碳布相比,MnO2修饰对电活性菌的固着能力增强,使其能够持续发挥电子桥梁作用。与传统碳布电极仅依靠阳极电势富集电活性菌群不同,MnO2凭借其优异的赝电容特性与可逆氧化还原特性,使Geobacter sp.稳定在更高丰度水平。高丰度的Geobacter sp.既保障了生物膜的双向电子传递功能,其含有的细胞色素C、导电菌毛及胞外电子中介体等组分,还强化了电极向反硝化菌的定向电子转移效率,并实现了在阳极期储存更多电子、在阴极期按需释放的功能。
菌群变化结果与前述电子平衡分析结果高度吻合,MnO2诱导Geobacter sp.大量富集,使得MnO2修饰组生物膜最大电子储存量高于空白组,同时,电极供给电子与生物膜储存电子总量均明显提升。菌群结构优化为电子储存与双向传递提供了微生物基础,而电子储存与释放能力的增强又进一步巩固了Geobacter sp.等核心功能菌的生态位优势。由此可见,MnO2修饰在传统“电活性菌—反硝化菌”协同脱氮体系之上,进一步构建了以高丰度Geobacter sp.为核心、兼具高效电子传递与生物赝电容特性的强化体系,使生物膜不仅能够高效传递电子,还可稳定存储与释放电子,从而在周期性极性反转条件下实现更持续、更高效的硝酸盐还原。
氮代谢功能基因预测结果进一步揭示了MnO2修饰提升脱氮效果的生物学机制(图 4b)。基于FAPROTAX功能预测结果,硝酸盐异化还原成铵、亚硝酸盐异化还原成铵相关功能基因的相对表达量在空白组与MnO2修饰组中均处于较低水平,且组间无明显差异,这与硝酸盐加入后期,系统出水仅有极少量氨氮产生的实验结果一致。MnO2修饰组中反硝化全通路基因呈现上调特征,硝酸盐还原、亚硝酸盐还原、一氧化二氮还原及总反硝化功能基因相对表达量均提升了2.17倍,激活了从硝酸盐→亚硝酸盐→一氧化氮→一氧化二氮→氮气的完整反硝化通路[27],且各步骤基因表达同步上调,避免了中间产物积累。Geobacter sp.的富集与反硝化基因上调形成协同作用,Geobacter sp.通过高效胞外电子传递系统为反硝化提供充足电子,反硝化基因的上调则增强了反硝化菌对电子的利用效率,形成“电子供体供给—电子受体利用”的协同,促进反硝化反应高效运行。综上所述,MnO2修饰通过富集电活性菌强化了反硝化电子供体供给,通过上调氮代谢功能基因激活了反硝化全通路,微生物群落演替与功能基因表达的协同调控共同缓解了低C/N废水反硝化能力不足的问题。
-
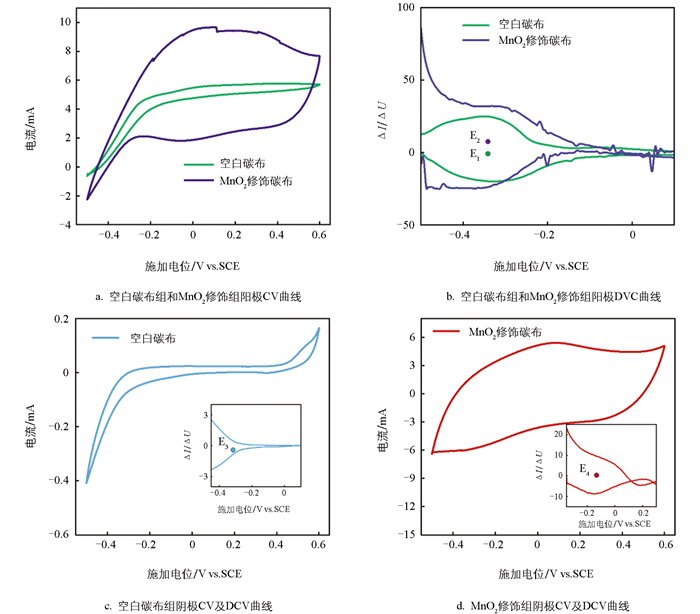
探究生物膜电化学活性位点对理解代谢过程及电子传递机制具有重要意义。如图 5a所示,在有底物状态下的循环伏安(CV)扫描测试中,阳极状态下CV曲线呈明显“S型”,这是以Geobacter sp.为优势菌的电活性生物膜的典型特征[28],表明阳极生物膜中富集了大量Geobacter sp.,这与微生物群落组成结果一致。MnO2修饰组表现出更强的电容特性,也印证了其在阳极状态能够储存更多电量的能力。由图 5b可知,阳极态生物膜的电化学活性位点均位于-0.34 V(E1、E2),这一电位与以往报道的以Geobacter sp.为主的生物膜电位特征一致[29]。在反硝化态下,有底物CV曲线与阳极态呈现显著差异,表明其群落结构发生演替。DCV曲线显示,空白碳布组和MnO2修饰组的电活性生物膜活性位点分别位于-0.31 V(E3) 以及-0.15 V(E4)。其中,-0.15 V处的活性位点推测主要归因于细胞色素C的低电位血红素中心[30]。在Geobacter sulfurreducens中,内膜多血红素细胞色素中点电位约为-0.15 V,负责将电子从内膜醌库传递至周质蛋白[31]。
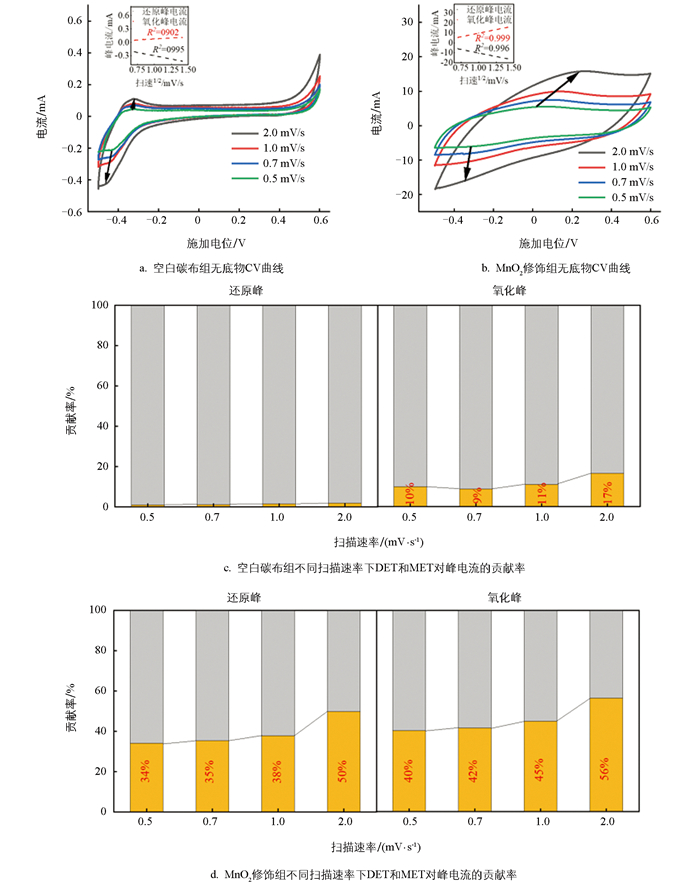
无底物状态下的CV测试是表征电极—生物膜电子传递过程的重要研究手段,图 6a和图 6b分别为空白碳布组和MnO2修饰组在无底物状态下的CV曲线,以及峰电流与扫描速率平方根之间的线性拟合结果。从电流密度绝对值来看,MnO2修饰组生物膜电流密度显著高于空白碳布组,表明MnO2修饰可显著提高生物膜导电性,增强电子转移能力,这与前述分析MnO2诱导生物膜产生更多细胞色素C或导电菌毛的分析结论一致。当扫描速率从0.5 mV/s增加至1.0 mV/s时,峰电流与扫速平方根呈显著正相关,表明此扫描速率范围内,反应主要受扩散过程限制,电子传递过程以间接电子传递为主。为精准反映直接电子传递(DET)与间接电子传递(MET)对电流的具体贡献,通过1.3节的公式计算二者对峰电流的贡献率。
图 6c和图 6d分别为空白碳布组和MnO2修饰组DET及MET对峰电流的贡献率。空白碳布组中,当扫描速率较小时(0.5 mV/s),还原态峰电流中MET贡献率接近100%,氧化态峰电流中DET贡献率为10%。随着扫描速率的增加,DET的贡献率有所增加,但无论是还原态还是氧化态峰电流中,DET贡献率仍未超过20%,说明空白碳布组电极-生物膜界面反应主要由扩散反应控制,电子传递过程以MET为主导。对MnO2修饰组还原态和氧化态峰电流的DET和MET贡献率进行计算发现,同样扫描速率下,其DET贡献率高于空白碳布组;但当扫速低于1.0 mV/s时,DET的贡献率仍低于50%,说明此时界面反应仍由扩散反应控制。当扫描速率提高至2.0 mV/s时,氧化态峰电流中DET的贡献率达到56%,即氧化态电流主要由动力学过程控制,电子传递以DET为主导,表明该体系具有更快的储存电子释放能力。DET贡献比例越高,表明生物膜-电极界面电子交换速率越快。DET主要与膜结合型细胞色素C、导电菌毛等直接相关,这些组分可作为生物膜与电极间的“电子桥”,实现电子的直接传递[32-33]。该结果与2.1节讨论中电子可能储存在细胞色素C中相吻合。在反硝化阶段,MnO2修饰组生物膜中的细胞色素C可作为“电子库”,释放其储存的大量电子,用于硝酸盐的还原。
-
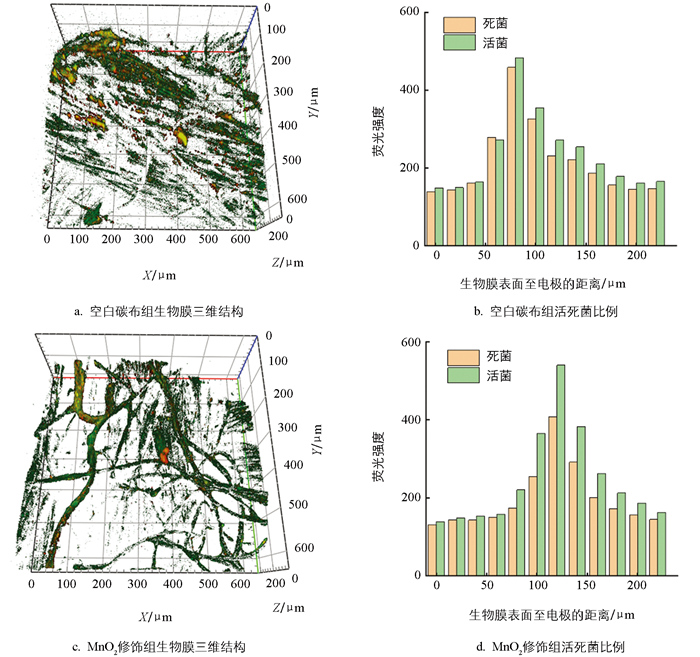
电极经修饰后,生物膜结构也随之发生变化。如图 7所示,空白碳布组与MnO2修饰组电极生物膜厚度均约为220 μm,但活菌空间分布存在显著差异。空白碳布组活菌荧光强度峰值出现在距电极80 μm处,MnO2修饰组则迁移至120 μm处,且生物膜表层(100~150 μm)活菌比例显著提升。结合本体系以硝酸盐氮为电子受体的特性,这种活菌空间分布差异对硝酸盐氮去除具有积极作用。
硝酸盐氮作为可溶性电子受体,通过扩散作用从溶液主体进入生物膜内部。空白碳布组活菌集中于近电极(80 μm)区域,硝酸盐氮需克服更大的扩散阻力才能到达活菌富集区,易形成表层硝酸盐氮浓度高、内部底物匮乏的传质瓶颈,导致部分硝酸盐氮未被充分降解便随溶液流出。而MnO2修饰组活菌向表层(120 μm)迁移后,活菌富集区与硝酸盐氮扩散前沿直接重合,大幅缩短了硝酸盐氮的扩散距离,减少了传质阻力,使溶液中的硝酸盐氮能快速与高活性活菌接触,提升了底物与微生物的作用效率。结合前文硝酸盐氮还原速率提升、中间产物积累减少的结果,可进一步证实MnO2修饰组的生物膜传质效率得到了有效优化。这种活菌“外移”现象,表明微生物在适应硝酸盐冲击时,可主动调节其在生物膜中的空间位置,而这一适应性变化与MnO2修饰强化了生物膜的电子传递能力密切相关。该体系中,反硝化过程是电子传递驱动的还原反应,电子传递效率直接决定硝酸盐氮的还原速率与效能。无底物状态下CV结果显示,MnO2修饰组的生物膜电流密度远高于空白碳布组,说明其导电性显著高于空白碳布组,这可能与MnO2诱导下生物膜大量分泌细胞色素C及导电菌毛等相关,进而搭建起高效的电子传输网络。空白碳布组活菌集中于近电极区,虽能直接获取电子,但电子需经多层生物膜传递至表层,受限于较差的生物膜导电性,表层硝酸盐氮无法获得充足电子,易出现硝酸盐还原不彻底的现象;而MnO2修饰组活菌外移后,依赖生物膜优异的导电性,保障了电子快速传输,使表层活菌快速获得来自电极的电子,为硝酸盐氮的多步还原反应提供了充足电子驱动力,提升了硝酸盐氮的还原速率,减少了中间产物的积累。
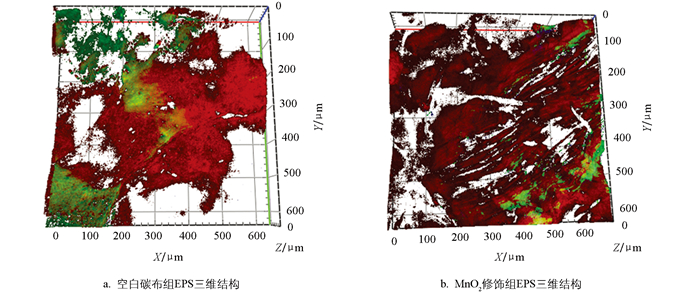
图 8为生物膜胞外聚合物(EPS)的激光共聚焦扫描显微成像及三维重构结果,其中,红色、绿色、蓝色分别对应EPS中的α-多糖、蛋白类物质和β-多糖。测试结果显示,空白碳布组EPS整体组成以α-多糖为主,β-多糖含量极低。α-多糖虽覆盖生物膜全域,但分布不均,呈块状聚集特征。MnO2修饰组α-多糖分布连续且均匀,形成三维网络骨架,为细菌黏附提供稳定支撑,同时蛋白类物质和β-多糖填充于α-多糖网络间隙,使EPS组成更均衡,表明生物膜结构稳定性得到了显著提升。连续的EPS三维网络可维持生物膜内部的水分平衡与菌群生理活性,确保系统在长期运行中仍能保持高效的硝酸盐氮去除性能,进一步提升了系统的稳定性。
2.1. 微生物电化学系统脱氮效能
2.2. 群落结构分析
2.3. 电极生物膜电化学特性解析
2.4. 生物膜三维结构分析
-
MnO2修饰碳布电极结合周期极性反转策略,可显著提升微生物电化学系统(MES)的反硝化脱氮效能,该修饰通过优化电极电化学特性、定向调控微生物群落结构、改善电活性生物膜形貌与组成,实现了电极-生物膜电子传递效率、反硝化功能基因表达及生物膜传质效率的协同提升。反应器阴极期起始电流密度达-10.2 A/m2,生物膜在阳极期的最大储电子量提升至(0.80±0.03) mmol,总氮去除率从初始36%提升至91%,且反应全程无亚硝酸盐积累。MnO2通过发挥自身电容特性,增强了生物膜的电子储存与释放能力,加速了硝酸盐还原;同时促使活菌向生物膜表层迁移,缩短了硝酸盐扩散距离,为反硝化营造适宜微环境,保障了系统脱氮效能的长期稳定。