


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
在西南紫色土区,植物根在防治坡面水土流失过程中扮演着十分重要的角色[1]。由于紫色土本身土层浅薄,且成土母质疏松、物理风化强烈、抗蚀性较弱[2],在降水集中、地形起伏较大等因素影响下,水土流失问题较为突出[3-4]。植物根可以防治土壤侵蚀,减少水土流失,起到固土护坡的作用[5]。植被根与土体结合形成根-土复合体[6],犹如天然的土壤加固器;同时,植被茎叶能够拦截降雨,减少雨滴对土壤表面的溅蚀,从而降低坡面径流速度,减少土壤侵蚀量[7]。植被根通过与土壤之间的相互作用,将土体所承受的剪应力转化为自身的拉应力,从而增强根-土复合体的稳定性[8-9]。因此当根受到外力作用时,其抗拉性能直接影响到根的固土能力[10]。
多种因素影响植物根的抗拉性能[11]。研究表明,标距长度会显著影响草本植物根的生物力学特性,其抗拉强度和断裂处的拉伸应变会随标距长度的增加而减小[12]。根的生物力学特性也跟树龄存在关联,根的抗拉强度和弹性模量会随着植株年龄的增长而提升[13]。此外,根中纤维素等化学物质的含量及其生理特性对拉伸性能有着显著影响[14-16]。总体而言,植物根的单根抗拉力学性能受根的形态、含水量、发育程度、组织成分等多种因素影响[11, 17-19]。西南地区降雨集中且强度较大,一方面易造成土壤颗粒流失及土壤含水率升高,减小根土之间的作用力,降低根的固土效果[20-22],增加坡面土壤的滑移和崩塌风险;另一方面,降雨引发的土壤含水变化,会间接影响根系自身的含水状态,而根系含水状态直接关联其核心力学性能,力学性能对于植物发挥固土护坡作用至关重要。鉴于此,本研究以香根草(Chrysopogon zizanioides)为研究对象,在重庆市万州区三峡香根草产业基地开展试验,采集香根草根,深入研究香根草根含水状态对其抗拉性能的影响,以期预测不同气候条件下紫色土区草本植物固土能力的差异,为减少水土流失提供理论依据。
全文HTML
-
试验区位于重庆市万州区三峡香根草产业基地(30°46′28″N,108°11′28″E),其海拔为1 664~1 710 m。此地年平均气温16.5 ℃,年平均降水量1 183.1 mm,降水主要集中在5月-9月,年平均日照时长1 168.1 h。试验区内土壤类型为中性紫色土,土壤质地以粉壤土为主,厚度为48~60 cm。
-
香根草作为一种多年生草本植物,其茎叶密集丛生,根系发达,须根密集且扎根深,能深入土壤数米,与土体结合形成根-土复合体[4]。香根草因其出色的水土保持和生态修复能力,已经被广泛应用于坡面水土流失治理工程中[20],在紫色土坡地的生态防护中发挥着重要作用。试验区内香根草于2020年5月种植,2022年6月进行野外调查及样品采集工作。试验设置3种根含水状态:自然含水状态(自然根)、饱和含水状态(饱和根)、风干状态(风干根)。
-
根样品采集:建立3个5 m×5 m的香根草种植样地,测量各株株高和冠幅后,选取各个样地内最接近平均值的3株(共9株)进行全根挖掘。根样品经保鲜袋封装后放置在冰箱4 ℃恒温保存。
土壤样品采集:在上述各样地及选定的无香根草种植的裸土对照样地共4个样地内,按照“S”形各布设5个样点,清理表层杂物及植株地上部分后,每个样点采集1个容重环刀(体积100 cm3),另外采集这5个样点的混合散土2 kg装袋。共计采集20个容重环刀以及4袋散土样。
-
采用环刀法测定土壤容重,采用筛分法测定土壤粒径分布,采用重铬酸钾外加热法测定土壤有机碳含量。采样地土壤的基本理化性质如表 1所示。
-
试验挑选外观顺直、直径均匀且表皮完整无损伤的根,用精度0.01 mm的数显游标卡尺在根两端及中间位置各测量1次直径,将3个点位的测量值取平均值作为该单根的直径d。根据直径大小把供试根划分为5个径级:径级Ⅰ(0 mm<d≤0.5 mm)、径级Ⅱ(0.5 mm<d≤1.0 mm)、径级Ⅲ(1.0 mm<d≤1.5 mm)、径级Ⅳ(1.5 mm<d≤2.0 mm)、径级Ⅴ(2.0 mm<d≤2.5 mm)。本试验设置了3种根含水状态处理:自然根处理,采回实验室后置于4 ℃冰箱内低温保鲜;饱和根处理,在试验前12 h用纯水浸泡,直至根质量不再增加,即已达水分饱和状态;风干根处理,在试验前于实验室阴凉通风处风干6 h。处理完毕的根在拉伸前,全部放置于保鲜盒中,以降低根水分的蒸发损耗。每种根含水状态处理下,每个径级挑选10根单根,截取11 cm用于拉伸试验,共计制备150根单根样品。
所有试验完成后,立即将根放入65 ℃烘箱内烘干48 h,取出后测定根干质量,并通过计算得出3种处理的单根含水量分别为31.57%、54.65%、16.99%。
-
试验开始前,在上述均匀且顺直的单根两端1.5 cm处均匀涂抹快干有机硅树脂,制得标距为80 mm的抗拉试验样本。采用微机控制电子万能试验机(CMT5105)测定单根极限抗拉力、极限抗拉应变,计算根各抗拉力学指标。
根抗拉强度是指植物根抵抗拉伸破坏的能力,通常以单位横截面积上所能承受的最大拉力来表示,计算公式为:
式中:T为根系的抗拉强度(MPa);Fm为单根极限抗拉力(N);d为根系直径(mm)。
根系极限延伸率是根系在受力拉伸过程中,从原始长度到断裂时的最大长度变化与原始长度之比,通常以百分比表示,计算公式为:
式中:δ为极限延伸率,以百分数(%)表示;L1是根系拉断时标距部分的长度(mm);L0是根系拉伸试验前的原始标距长度(mm)。
根系弹性模量也叫杨氏模量,计算公式为:
式中:E为弹性模量(MPa);Δσ为弹性阶段内应力变化量(MPa);Δε为弹性阶段内应变变化量(%)。
-
本研究使用Excel 2019进行数据处理,Origin 2021进行数据拟合及绘图,SPSS 24.0进行ANOVA单因素方差分析和双因素方差分析。
1.1. 试验区概况
1.2. 试验设计
1.3. 样品采集
1.4. 试验方法
1.4.1. 土壤理化性质
1.4.2. 根系直径及含水状态
1.4.3. 根系单根抗拉力学特性
1.5. 数据处理及分析
-
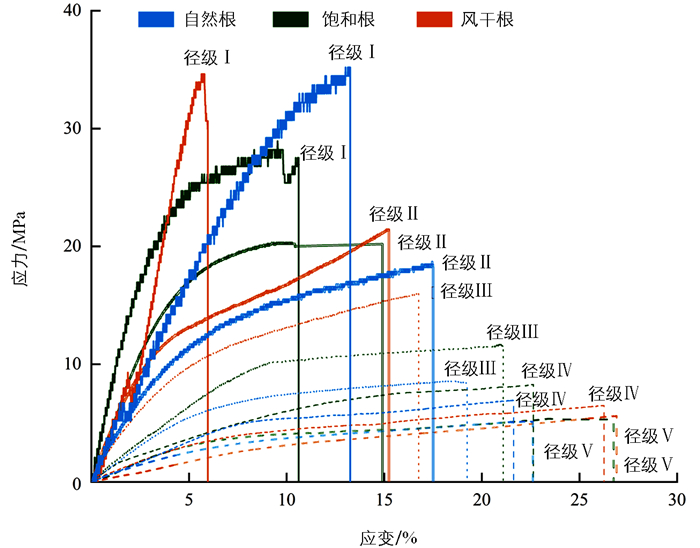
分别在5个径级的香根草根中各选取1根具有代表性直径的单根制作应力-应变曲线(图 1)。不同含水状态处理下根各径级的曲线表明:径级Ⅱ、Ⅲ、Ⅳ、Ⅴ的曲线呈现出相同趋势,即受力初期,应力随应变的增加呈线性增加,根表现出弹性变形,随后单根荷载超过弹性极限,应力-应变曲线斜率逐渐减小,即单根从弹性阶段进入塑性变形阶段;径级Ⅰ的应力-应变曲线存在差异,饱和根、自然根径级Ⅰ的曲线都存在弹性变形和塑性变形两个阶段,但饱和根的塑性变形阶段更长,风干根则在断裂前仅表现出弹性变形;各含水状态处理下径级Ⅰ、Ⅱ的应力-应变曲线斜率明显大于径级Ⅲ、Ⅳ、Ⅴ,表明在相同伸长量时,较细根单位面积上所能承受的拉力更大,随着拉伸应变的增加,应力迅速增大。
-
不同含水状态处理下根抗拉强度的研究结果(表 2)显示:随着含水量的增加,香根草各径级根的抗拉强度均逐渐降低;风干根的抗拉强度与自然根的差异无统计学意义;径级Ⅰ、Ⅱ、Ⅲ饱和根的抗拉强度较自然根分别显著降低了12.44%、34.10%、11.20%,而径级Ⅳ、Ⅴ饱和根的抗拉强度较自然根的差异无统计学意义。
在各径级当中,不同含水状态处理的香根草单根极限延伸率间的差异无统计学意义,但含水状态对不同径级根极限延伸率的影响不同。径级Ⅰ、Ⅱ的单根极限延伸率会随含水量增加而减小;而径级Ⅲ、Ⅳ的单根极限延伸率则会随含水量增加而增大。
香根草饱和含水状态下径级Ⅲ根的弹性模量较自然含水状态显著降低了28.47%,其余径级均没有发生显著性变化。风干状态下大径级根(径级Ⅲ、Ⅳ、Ⅴ)的弹性模量较自然含水状态分别显著提高了34.18%、25.10%、35.92%,其余径级均没有发生显著性变化。
-
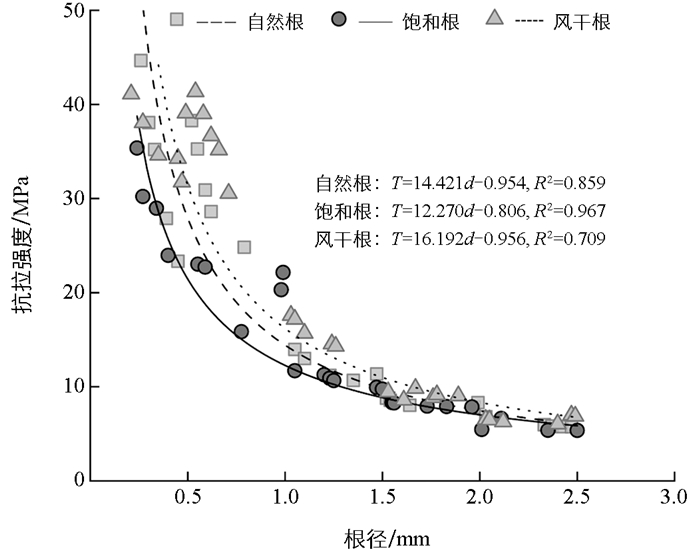
不同含水状态下,香根草单根的抗拉强度均随着根径增大而降低,根径越大,抗拉强度降低的幅度越小(图 2)。3种含水状态处理下,根径与单根抗拉强度的拟合方程决定系数R2分别为0.859、0.967、0.709,表明幂函数模型能很好地拟合两者间的关系。通过双因素方差分析可知(表 3),根含水状态、根径对单根抗拉强度都有极显著影响(F=21.44,p<0.01;F=223.87,p<0.01),且根含水状态和根径对单根抗拉强度有极显著交互作用(F=4.80,p<0.01)。通过单因素方差分析(表 4),检验在不同含水状态下,各径级间单根抗拉强度的差异是否有统计学意义,结果表明3种含水状态下,根径对单根抗拉强度都有极显著影响(p<0.01),其中F为交互作用值。
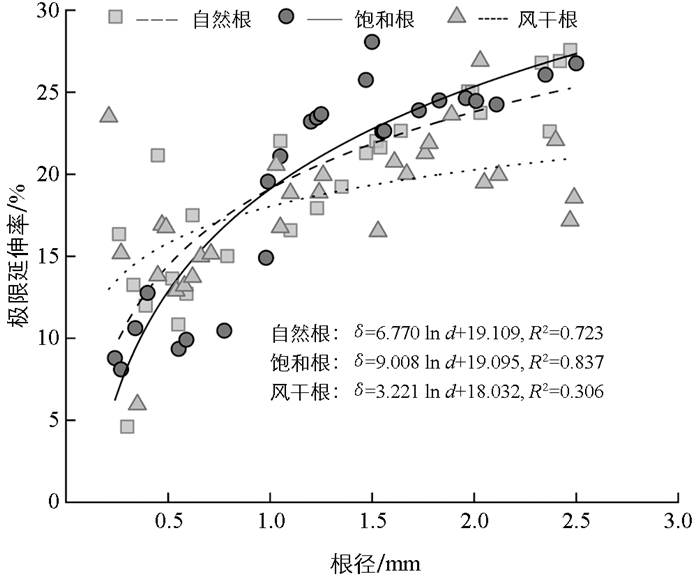
由图 3可知,在不同含水状态处理下,香根草单根极限延伸率均随着根径的增大而增大,两者关系可以用对数函数进行估计。但风干状态下的回归函数预测精度相对较低,拟合方程决定系数R2仅为0.306。通过根含水状态和根径与单根抗拉力学性能的双因素方差分析可知(表 3),根径对单根极限延伸率影响极显著(F=39.71,p<0.01),而根含水状态对单根极限延伸率影响不显著(F=1.37,p=0.26),根含水状态和根径对单根极限延伸率有极显著交互作用(F=3.09,p<0.01)。通过单因素方差分析(表 4),对不同含水状态下各径级间单根极限延伸率进行显著性检验,结果表明自然根、饱和根的根径会显著影响单根的极限延伸率(p<0.01),而风干根的单根极限延伸率受根径的影响并不显著(p=0.25)。
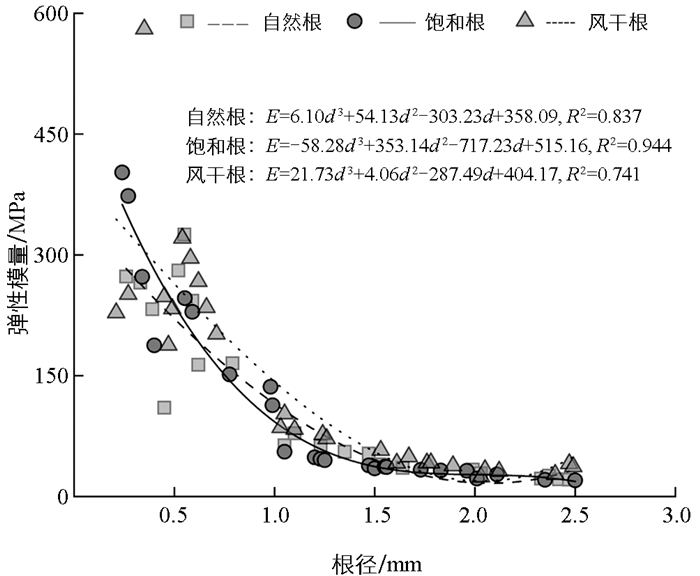
由图 4可知,在不同含水状态下,香根草单根弹性模量均随着根径的增大而降低,根径越大,弹性模量降低的幅度越小。根径与弹性模量的拟合方程决定系数R2分别为0.837、0.944、0.741,表明三次函数模型能很好地拟合两者间的关系。通过双因素方差分析可知(表 3),根径对单根弹性模量有极显著影响(F=31.16,p<0.01),而根含水状态对单根弹性模量影响不显著(F=0.61,p=0.55)。通过单因素方差分析(表 4),结果表明3种含水状态下,根径对单根弹性模量都有极显著影响(p<0.01)。
2.1. 根拉伸试验的应力-应变曲线
2.2. 单根抗拉强度、极限延伸率和弹性模量
2.3. 不同含水状态下根径对抗拉性能指标的影响
-
根的弹塑性变形指根在外力(如水流冲刷、重力)作用下,既能发生可恢复的弹性变形,也能产生不可恢复的塑性变形,这种特性受土壤含水量影响显著。研究发现,香根草根饱和含水状态会延长径级Ⅰ(0 mm<d≤0.5 mm)单根的塑性变形阶段,而香根草根风干状态会导致径级Ⅰ单根在拉伸至断裂过程中只发生弹性变形而没有明显的塑性变形阶段。这是由于径级Ⅰ的单根属于香根草的幼嫩细根,细胞壁以初生壁为主,木质化程度低,细胞间隙较大,高含水状态下更易发生变形,且细胞内原生质体对水分变化敏感,水分状态直接影响细胞的膨压性和细胞壁的延展性。香根草径级Ⅰ根的塑性变形本质上是细胞壁延展性、细胞间连接状态及原生质体黏弹性共同作用的结果,而水分是调控这些特性的关键因素。饱和含水状态处理通过增强细胞壁延展性缓冲细胞间应力,延长塑性变形阶段。风干状态处理通过强化细胞壁刚性破坏细胞整体性,导致塑性阶段消失,仅存弹性变形直至断裂。Boldrin等[23]的研究结果也表明根应力-应变曲线的弹塑性受风干状态处理的影响显著。但Yang等[24]关于根应力-应变曲线的研究显示,大径级根风干后在拉伸过程中也只发生弹性变形直至根断裂。这可能是因为其研究所采用的是蒙古栎、油松等乔木的根,这些大根细胞壁中木质素含量较高[25],增强了细胞壁的刚性和硬度,减少了塑性变形的可能性[26]。
-
本研究结果发现香根草单根抗拉强度会随着含水量的增加而减小,这与Mahannopkul等[27]关于根含水量与抗拉性能的研究结果一致。这是因为细胞大量吸水而膨胀,导致细胞壁强度降低,而细胞壁是影响根抗拉力的关键结构,其强度下降,单根抗拉强度也会减小[20]。此外,有研究发现根含水量变化对单根抗拉强度的影响程度会随着根直径的增加而下降[28],这是因为较大直径根有更复杂丰富的组织结构,能缓冲薄壁细胞受含水量变化的影响[29-30],使整体抗拉强度受影响小。本研究还发现,饱和含水状态会显著降低径级Ⅰ、Ⅱ、Ⅲ根的抗拉强度,而风干状态对根抗拉强度无显著影响。这可能是因为香根草根在风干过程中,虽然水分大量流失,但木质素等抗降解能力较强的物质并没有因风干处理而发生显著变化[30],使得风干处理对其整体抗拉强度的影响不明显。任何含水状态下香根草根径对单根抗拉强度有极显著影响,这与肖海等[10]的研究结果相似。
不同径级中香根草根含水状态对单根极限延伸率的影响不同,且根含水状态不会对根的单根极限延伸率水平造成显著影响。这可能是因为较细的根(d≤1.0 mm)由于薄壁细胞本身的结构相对脆弱,过度膨胀可能使细胞壁的结构受损,导致单根极限延伸率随含水量增加而减小。而1.0 mm<d≤2.5 mm的根中具有一定比例的厚壁组织、木质部等结构[31],使得根在拉伸时既能发生较大的变形,又能保持结构的完整性,从而单根极限延伸率随含水量增加而增大。有研究认为根极限延伸率随根径的增加而增大[32],也有研究显示根极限延伸率随根径的增加呈现减小的趋势[33],上述研究均认为根径不会对单根极限延伸率造成显著影响。但本研究结果显示根径对单根极限延伸率有显著性影响,且根含水状态和根径对单根极限延伸率有极显著的交互作用。这可能是因为不同研究以不同植物为对象,不同植物间的根结构和生理特性存在差异,导致在拉伸过程中植物根极限延伸率随根径的变化呈现出不同的增减趋势,以及根径对单根极限延伸率的影响程度存在差异。在自然含水状态、饱和含水状态下根径会显著影响单根的极限延伸率,风干状态下根的单根极限延伸率受根径的影响并不显著,表明根含水状态会影响根径与单根极限延伸率之间的关系。
除径级Ⅰ的根外,径级Ⅱ、Ⅲ、Ⅳ、Ⅴ的根弹性模量均随含水量的增加而降低,与前人的研究结果相似。这是因为水分弱化了香根草根细胞壁的成分连接,松弛细胞结构并降低组织协同受力能力,导致其抵抗变形的能力下降。风干状态下,大径级根(径级Ⅲ、Ⅳ、Ⅴ)的弹性模量较自然含水状态显著提高,而其他径级根无显著变化,核心原因在于不同径级根的结构特性和水分依赖度差异。任何含水状态下香根草根径对单根弹性模量都有极显著影响,且随根径的增加,单根弹性模量降低,这与前人的研究结果相似。这是因为香根草小径级根细胞排列致密,纤维素含量高且排列整齐,弹性模量较高以适应吸收功能;而大径级根因次生生长导致结构疏松、木质素占比上升,加之其支撑与运输功能需要一定柔韧性,使得弹性模量降低。
-
1) 香根草根在不同含水状态下,其应力-应变曲线存在差异。香根草径级Ⅰ(0 mm<d≤0.5 mm)饱和根存在弹性变形与塑性变形,且塑性变形阶段较自然根更长。径级Ⅰ的风干根在拉伸过程中只发生弹性变形,没有明显的塑性变形阶段直至断裂。在各含水状态下,0 mm<d≤1.0 mm的根比1.0 mm<d≤2.5 mm的根弹性模量更大。表明香根草小径级根在高含水(饱和)状态下,其抗拉伸断裂能力更优,更有利于固持土壤。
2) 根含水状态变化对单根抗拉强度的影响程度随根径增加而下降。饱和含水状态会降低0 mm<d≤1.5 mm根的抗拉强度和1.0 mm<d≤1.5 mm根的弹性模量。风干状态会提升1.0 mm<d≤2.5 mm根的弹性模量。饱和含水状态下0 mm<d≤1.5 mm各径级单根抗拉强度较自然含水状态分别显著降低了12.44%、34.10%、11.20%;1.0 mm<d≤1.5 mm径级单根弹性模量较自然含水状态显著降低了28.47%。而风干状态下香根草1.0 mm<d≤2.5 mm各径级根的弹性模量分别较自然含水状态提高了34.18%、25.10%、35.92%。表明香根草大径级根在低含水(风干)状态下,其整体抗拉性能更优。
3) 根含水状态会影响根径与单根极限延伸率之间的关系。不同含水状态下,根径对单根抗拉强度、弹性模量都有极显著影响。自然含水状态、饱和含水状态下的根径会显著影响单根的极限延伸率,而风干状态下单根极限延伸率受根径的影响不显著。