


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
苹果炭疽叶枯病(Glomerella Leaf Spot,GLS)是由围小丛壳(Glomerella cingulata)侵染引起的一种真菌病害,其无性态为炭疽菌属(Colletotrichum)真菌[1-2]。该病害主要侵染叶片和果实,导致叶片焦枯脱落、果实褐斑腐烂,造成树势衰退、产量骤降及果实商品性丧失[3]。自1988年在巴西首次报道以来[4],GLS因潜伏期长、暴发性强、防治难度大、为害严重,已成为影响我国苹果产业可持续发展的主要真菌病害之一。目前主要依赖传统化学防治,长期使用易引致抗药性和农药残留问题。利用植物免疫诱抗剂诱导激活植物自身抗病性,已成为绿色防控的重要策略,对苹果产业可持续发展具有重要意义。
植物免疫诱抗剂,又称植物疫苗,是一类新型生物农药。该类物质虽无直接杀菌或抗病毒活性,但能通过激活植物先天免疫系统,实现抗病、增产的效果,同时有助于减少化学农药的使用量,符合绿色植保理念。植物免疫诱抗剂主要分为5大类:无机化合物类、有机酸类、苯并噻二唑类、寡糖类、蛋白多肽类和免疫诱抗菌类[5]。其中,寡糖类植物免疫诱导剂在我国研究应用较早。该类诱导剂的分子结构与真菌细胞壁的降解产物高度相似,能够模拟病原菌侵染过程,从而激活植物体内病程相关蛋白的表达,有效诱导植物系统抗病性的形成[6]。香菇多糖(Lentinan,LNT)作为代表性寡糖类诱抗剂,是一种从香菇(Lentinus edodes)子实体或菌丝体中提取的天然β-葡聚糖,对植物、人畜和环境无害,符合绿色农业需求[7]。自1968年被日本学者千原吴郎首次分离以来,LNT除作为肿瘤辅助治疗药物外[8],在植物保护领域也显示出良好的应用潜力。单宏英等[9]研究发现1% LNT溶液对烟草灰霉病的防治效果达85.08%;王胤等[10]发现其可显著抑制番茄灰霉病菌菌丝生长。在抗病毒方面,Wang等[11]研究表明LNT通过诱导H2O2暴发及上调PR2、Chitinase、PAL等抗病基因表达,抑制烟草花叶病毒(TMV)侵染与复制;鄢兴祥[12]试验显示10 g/L LNT对番木瓜花叶病毒病防效为82.7%,并能提高番木瓜的株高和茎粗;姜莉莉等[13]发现LNT与化学杀菌剂混用可协同防治小麦纹枯病和根腐病,实现减药控害目标。因此,LNT作为一种高效安全的植物免疫诱抗剂,兼具抗病、促生和化学农药减量增效功能。然而,目前有关LNT对GLS防控效果的研究较少,其作用机制和实际应用价值尚待研究。
本研究拟以“嘎啦”苹果组培生根苗为试材,通过设置不同浓度的LNT喷施处理,结合含药平板抑菌与离体叶片接种试验,筛选出可诱导苹果植株产生最大GLS抗病性的最适LNT稀释浓度。在最佳浓度处理下,通过组织学方法观察LNT对病原菌侵染进程的抑制作用,并系统测定活性氧含量、抗氧化酶活性、内源激素水平、氨基酸和酚类次生代谢物的积累量,以及病程相关基因的表达动态,以初步解析LNT提高苹果对炭疽叶枯病抗病性的生理与分子机制。基于室内研究结果,进一步开展田间防控试验,评估LNT对苹果叶部病害的实际防治效果,以明确其农业应用潜力,为苹果病害的绿色防控提供理论依据与实践指导。
全文HTML
-
以“嘎啦-3”(GL-3)苹果组培苗为试材,经继代培养生根后移栽至营养钵。待幼苗长至20 cm高时,选取茎秆粗壮、木质化程度一致的植株用于试验。苹果炭疽叶枯病病原菌由青岛农业大学李保华教授馈赠,2%香菇多糖水剂购自山东圣鹏科技股份有限公司。
-
将GLS病原菌在PDA培养基上于25 ℃黑暗条件下培养,取5 mm菌饼转接至PDB液体培养基中,25 ℃、140 r/min振荡培养4 d,收集孢子并调配浓度至1×106 CFU/mL。接种时,用喷雾器将孢子悬浮液均匀喷施于叶片表面,以叶片正反面均被细密雾滴完全覆盖,但无液滴形成为宜,叶柄基部包裹湿棉条保湿。接种后植株置于(30±1) ℃、相对湿度(90±5)%、黑暗的人工气候箱培养。
-
LNT适宜浓度筛选:取60株苹果幼苗,随机分为6组(5个处理组和1个对照组),每组10株。将2% LNT水剂分别稀释200倍、400倍、600倍、800倍和1 000倍。分别对5个处理组叶面喷施上述对应浓度的LNT稀释液,每日2次,连续处理2 d;对照组喷施等量清水。每处理组取30片苹果叶片接种GLS病原菌,72 h后测定病斑面积和相对电导率(REL)等指标,以确定最佳诱抗浓度。
LNT诱抗效应分析:取80株幼苗,随机分4组,每组20株。① CK组(喷施清水,不接种GLS病原菌);② LNT组(喷施800倍LNT稀释液,不接种);③ CK+GLS组(喷施清水后接种);④ LNT+GLS组(喷施800倍LNT稀释液后接种)。每组取200片健康叶片,于接种后0、6、12、24、48和72 h取样,液氮速冻后-80 ℃保存,用于生理指标及基因表达分析。剩余叶片用于病斑面积统计和活性氧染色等组织化学观察。
-
分别将500、250、166、125和100 μL的2% LNT水剂加入45 ℃融化的PDA培养基中,定容至100 mL,配制成稀释倍数分别为200、400、600、800和1 000倍的含药平板培养基,以加等量无菌水的PDA平板为对照。每组设2次重复。取直径5 mm的GLS菌饼接种于平板中央,25 ℃黑暗培养5 d,十字交叉法测量菌落直径。计算菌丝生长抑制率:
-
使用EPSON perfection V700 photo扫描仪获取叶片图像,采用ImageJ Pro软件定量分析病斑面积。
-
分别于0 d和3 d时从各组采集幼嫩叶片3~5片,去离子水冲洗干净,吸干表面水分后测定相对电导率[14]。
-
取1.5 cm×1.5 cm叶片样品,于4 ℃下用FAA固定液固定24 h。经梯度乙醇脱水、二甲苯透明、石蜡包埋后,切成8~10 μm厚度的切片,贴片烘干。切片脱蜡复水,苯胺蓝染色,再经脱水透明,中性树脂封片,于奥林巴斯BX-53显微镜下观察拍照。
-
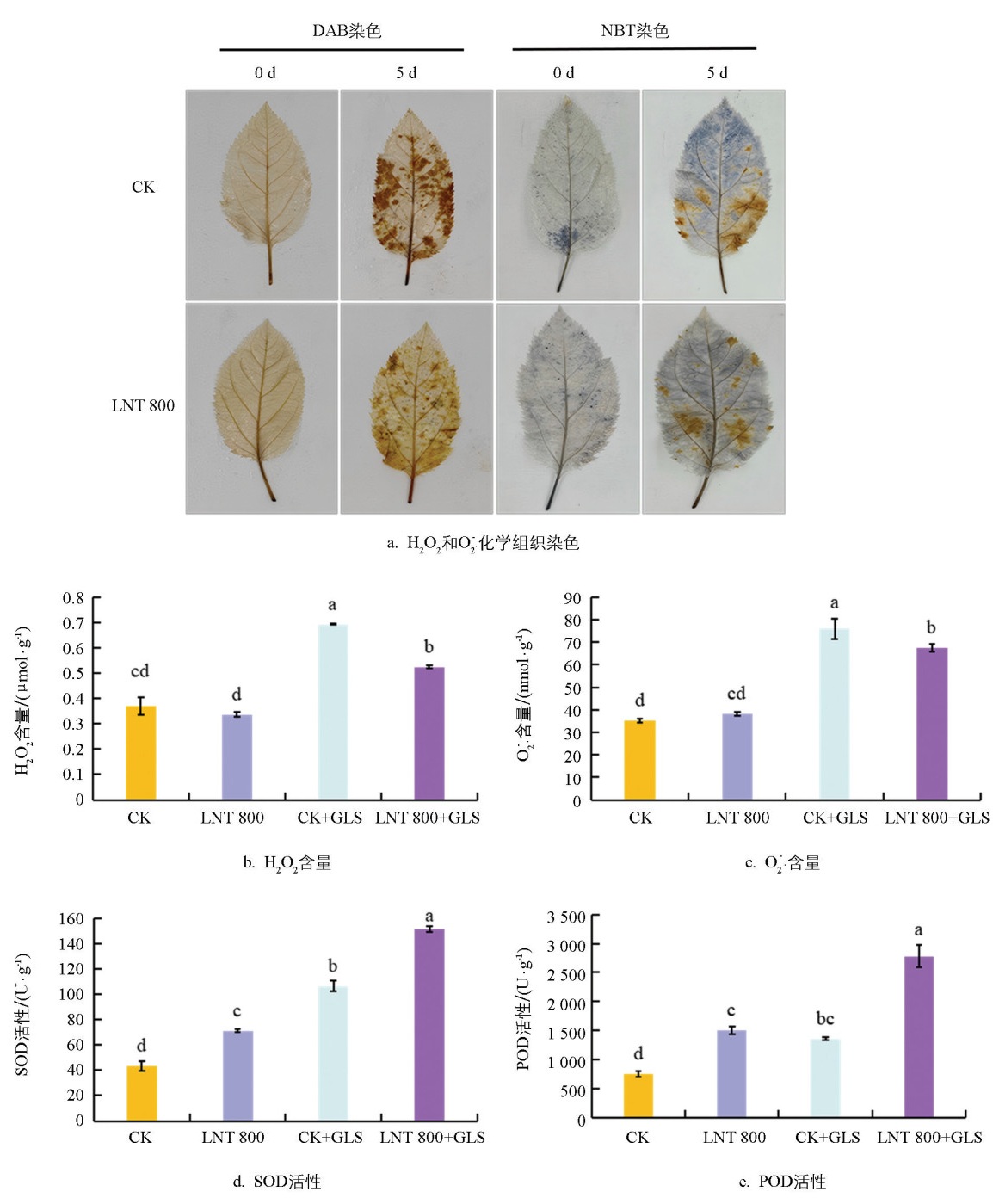
采用DAB染色检测H2O2(棕色),NBT染色检测O2·-(蓝色)。DAB染色液为含0.5% DAB和0.1% TritonX-100的PBS缓冲液,NBT染色液为含0.5 g/L NBT和0.1% TritonX-100的HEPES缓冲液(pH值7.5)。叶片分别浸入相应染色液,抽真空避光静置(DAB过夜,NBT 1 h)。染色后,用脱色液(乳酸∶甘油∶无水乙醇=1∶1∶3)沸水浴脱色至叶绿素完全褪去,制片镜检。
-
采用H2O2-2-Y、SA-2-G、SOD-2-Y和POD-2-Y试剂盒(苏州科铭生物技术有限公司)分别测定叶片中H2O2和O2·-含量以及超氧化物歧化酶(SOD)和过氧化物酶(POD)的酶活,操作按说明书进行。
-
提取氨基酸并进行对应的含量测定[15],检测包括脯氨酸、精氨酸、赖氨酸、苏氨酸、苯丙氨酸、酪氨酸、组氨酸、丝氨酸、丙氨酸、甲硫氨酸、缬氨酸、异亮氨酸、亮氨酸、甘氨酸、谷氨酸和天冬氨酸等16种氨基酸的含量。
-
提取酚类物质并进行含量测定[15],分别检测根皮苷、肉桂酸、儿茶素、原花青素、芦丁、槲皮素、金丝桃苷、异栎素、表儿茶素、对香豆酸、没食子酸、山柰酚、原儿茶酸、阿魏酸和绿原酸等15种酚类物质的含量。
-
提取激素并进行含量测定[16],检测包括玉米素(ZT)、生长素(IAA)、1-氨基环丙烷-1-羧酸(ACC)、赤霉素(GA3)、水杨酸(SA)、6-苄氨基嘌呤(6-BA)和脱落酸(ABA)等7种激素的含量。
-
于接种GLS后0 h、3 h、6 h、12 h、24 h和48 h取样,液氮速冻后-80 ℃保存。采用Fast Pure Universal Plant Total RNA Isolation Kit(Vazyme)试剂盒提取叶片总RNA,HiScript IV All-in-one Ultra RT Super Mix for qPCR试剂盒反转录合成cDNA。以cDNA为模板,使用表 1中PR1、PR2、PR4、PR5、PAL、PPO和Chitinase基因的引物进行qRT-PCR。反应程序:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火1 min,共30个循环。采用2-ΔΔCT方法[17]计算基因相对表达量。
-
将叶片切成1 cm×1 cm小块,用95%乙醇脱色至透明(每2 h更换脱色液)。以含0.01%苯胺蓝的150 mmol/L K2HPO4缓冲液(pH值9.5)避光染色过夜。叶片于50%甘油中保存并制片,在激发波长400~485 nm下观察胼胝质沉积。
-
田间试验于2024年在陕西省宝鸡市扶风县青龙庙村开展,设处理和对照:喷施800倍LNT稀释液,对照喷施等量清水。每处理120棵树,间隔一行果树以防交叉污染。5月10日首次施药,使用芝浦ZP-LDDYQ背负式电动静电喷雾器(工作压力0.2~0.4 MPa,流量1 L/min,雾滴中径≤120 μm)喷雾,以叶片润湿不滴液为施药标准。每15 d喷施1次,共3次,试验期间未使用其他杀菌剂。末次喷施后第7 d,测定株高、新梢数量、新梢长度和冠径等生长指标,并按照分级标准[18]调查发病情况、统计病级、计算病情指数和诱抗防治效果。
-
采用GraphPad Prism v10.1.2软件进行单因素方差分析(ANOVA)及Tukey's检验(p<0.05),数据以平均值±标准误(SE)表示。使用Origin 2019软件(OriginLab,Northampton,USA)绘图,TBtools软件绘制氨基酸、酚类物质和激素含量热图。
1.1. 试验材料
1.2. 试验方法
1.2.1. GLS病原菌活化与苹果离体叶片接种
1.2.2. 试验设计
1.2.3. LNT对GLS菌丝生长抑制效果的测定
1.2.4. 叶片病斑面积测定
1.2.5. 相对电导率测定
1.2.6. 叶片切片制作及显微结构观察
1.2.7. DAB和NBT组织化学染色
1.2.8. 活性氧含量及抗氧化系统相关酶活性的测定
1.2.9. 氨基酸的提取及含量测定
1.2.10. 酚类物质的提取及含量测定
1.2.11. 内源激素的提取及含量测定
1.2.12. 病程相关基因表达分析
1.2.13. 胼胝质沉积染色
1.2.14. 田间喷施LNT对苹果生长和叶部病害抗性诱导效应分析
1.2.15. 数据处理与分析
-
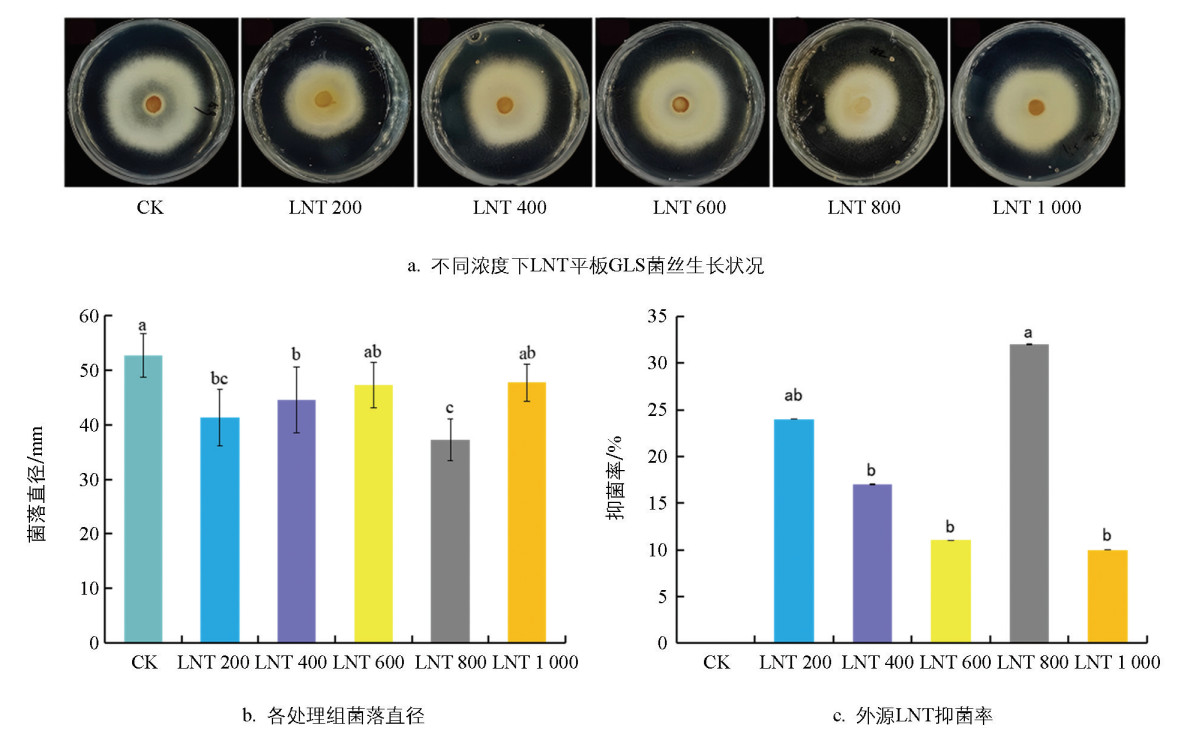
试验结果看出,CK组的菌落直径最大,600倍和1 000倍LNT稀释液处理组与CK组无显著差异,200倍和400倍处理组的菌落直径小于CK组,800倍处理组的菌落直径最小,为37.20 mm(图 1a、b)。抑菌率分析表明,800倍稀释液的抑菌率最高,为32.49%(图 1c),说明该浓度对GLS菌丝生长的抑制作用最强。
-
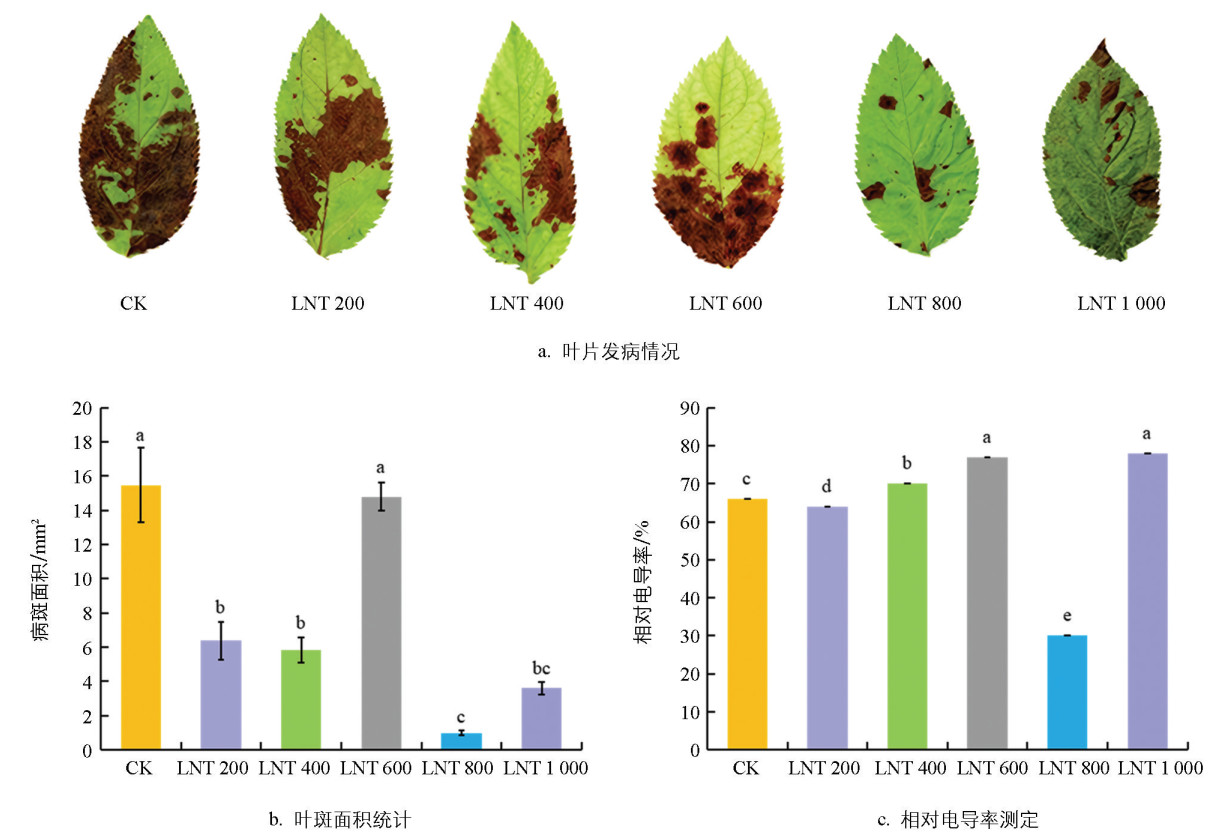
苹果离体叶片接种GLS后,200、400、600、800和1 000倍LNT稀释液处理组的病斑面积均显著小于CK组(图 2a),表明LNT能缓解病原菌造成的叶片组织坏死,其中800倍稀释液处理组的病斑面积最小,较CK组减小93.58%(图 2b)。接种3 d后,仅800倍处理组的相对电导率显著低于CK组,降至30.34%(图 2c)。以上结果表明,800倍LNT稀释液能有效抑制病斑扩展并减轻细胞膜损伤,后续试验选用该浓度。
-
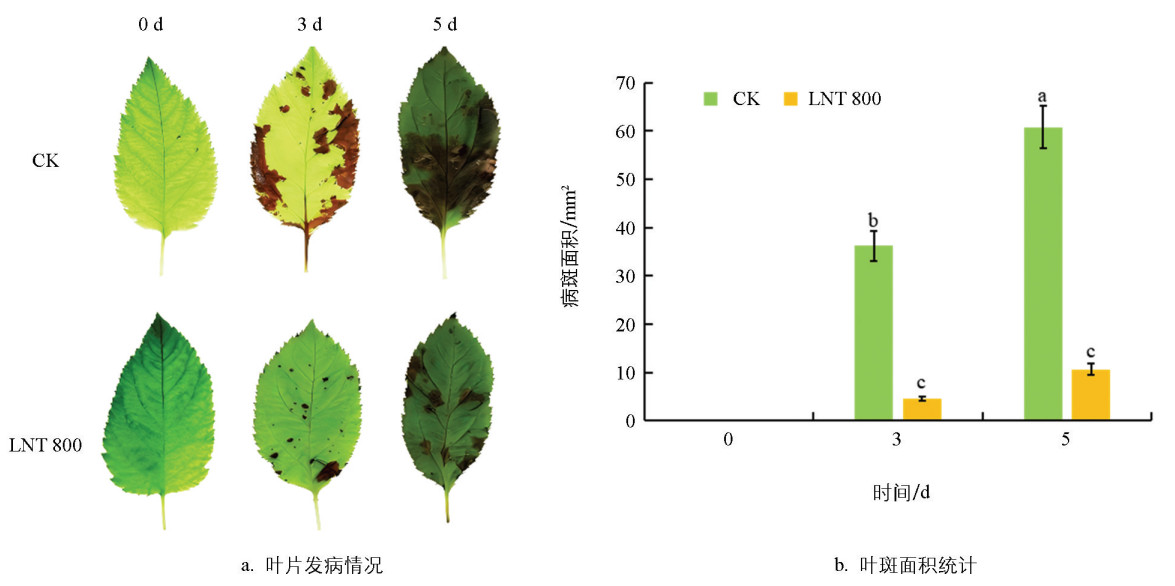
接种GLS后,CK组病斑面积从3 d时的36.30 mm2扩大至5 d时的60.80 mm2(图 3a)。相比之下,LNT处理组的病斑扩展显著受限,在3 d和5 d时的病斑面积分别为4.55 mm2和10.69 mm2,仅为同期CK组的12.54%和17.68%(图 3 b)。表明LNT处理可显著抑制GLS侵染引起的病斑扩展。
-
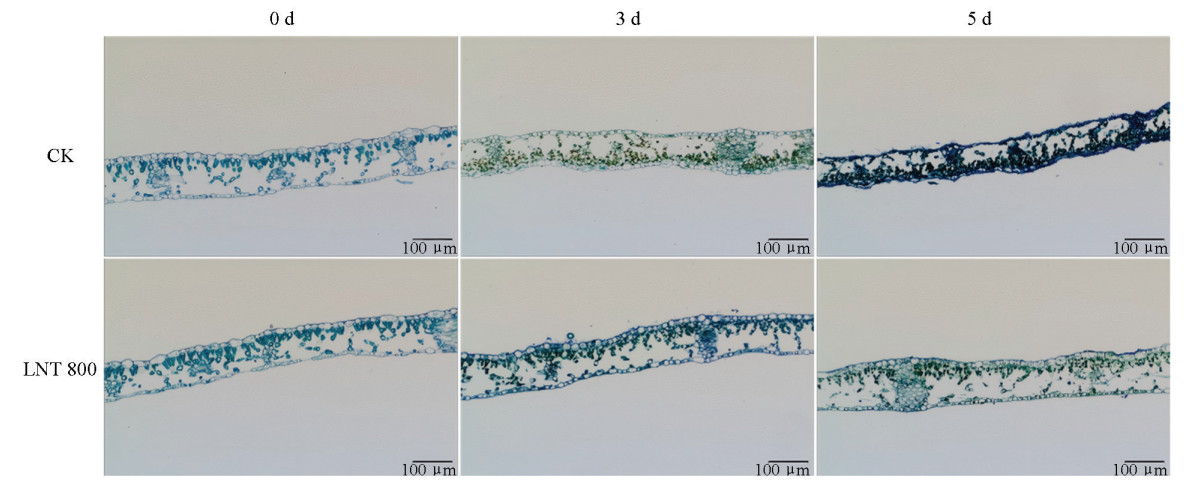
接种GLS后,CK组与LNT组叶片显微结构差异显著(图 4)。第0 d,叶片结构完整,细胞饱满。接种3 d后,CK组表皮细胞受损,栅栏组织和海绵组织褐变;LNT组仅轻微损伤,结构良好。接种5 d时,CK组结构严重崩解;LNT组损伤加重,但完整性相当于CK组第3 d水平,细胞仍可辨。表明LNT处理能延缓GLS引起的组织破坏,维持细胞完整性,减轻病害。
-
DAB和NBT染色显示,第0 d时两组叶片均无明显染色。接种GLS后,CK+GLS组叶片中H2O2(棕色)和O2·-(蓝色)大量积累,而LNT处理后两者显著减少(图 5a)。定量结果与染色一致:LNT组H2O2含量低于CK组,接种GLS后,CK+GLS组升高,而LNT+GLS组较CK+GLS组降低23.47%(图 5b)。O2·-含量变化趋势类似,LNT+GLS组较CK+GLS组降低11.11%(图 5c)。SOD活性以CK组最低(43.35 U/g),LNT处理后升高,接种GLS后,CK+GLS组进一步升高,LNT+GLS组达到最高值(151.54 U/g),显著高于CK+GLS组(图 5d)。POD活性变化趋势相同,LNT+GLS组最高,显著高于CK+GLS组(图 5e)。上述结果表明,LNT处理能增强SOD和POD活性,减少GLS侵染诱导的活性氧(ROS)积累,缓解氧化胁迫,提高苹果叶片抗病性。
-
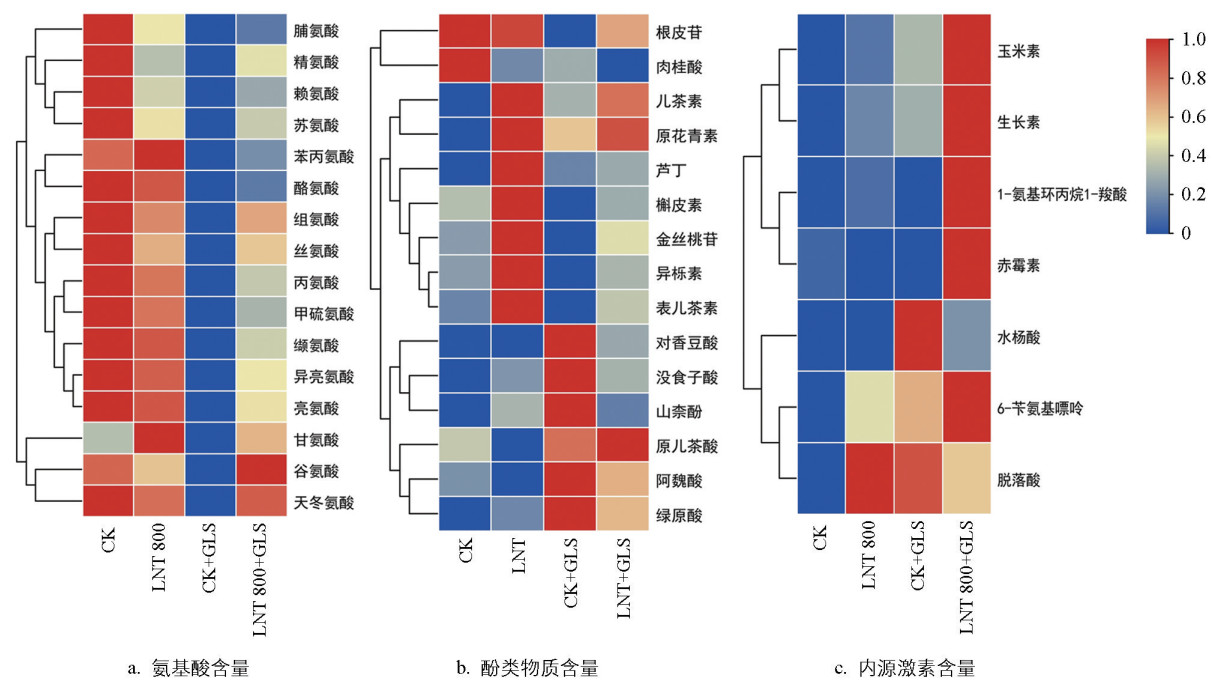
在氨基酸代谢方面(图 6a),与CK组相比,LNT处理使甘氨酸和苯丙氨酸含量升高了29%和12%,精氨酸等下降。CK+GLS组中多数氨基酸含量最低,其中脯氨酸和精氨酸分别仅为CK组的11.29%和17.09%。LNT+GLS组中各氨基酸含量均较CK+GLS组显著提高,以甲硫氨酸增幅最大(3.63倍)。聚类分析显示,脯氨酸和精氨酸在CK组中最高,接种GLS后降低,聚为一簇;苯丙氨酸和酪氨酸变化趋势相似,聚为另一簇;甘氨酸和谷氨酸变化不同,单独成簇。
在酚类物质代谢方面(图 6b),与CK相比,LNT处理使表儿茶素等8种酚类物质含量提高,芦丁增幅最大,提高了59%。CK+GLS组中原儿茶酸等6种酚类物质含量较CK组显著上升,以绿原酸(10.73倍)和对香豆酸(6.26倍)最为显著。LNT+GLS组中根皮苷等9种酚类物质较CK+GLS组进一步升高,其中异栎素和金丝桃苷增加最明显,分别提高了39%和31%,而绿原酸等4种酚类物质则下降至CK+GLS组的17.77%~66.10%。聚类分析显示,芦丁、金丝桃苷、槲皮素等在LNT组积累较多,对香豆酸和绿原酸等在CK+GLS组积累较高。
在激素调控方面(图 6c),未接种GLS时,LNT组中玉米素含量为0.02 ng/g(CK组未检出),脱落酸、6-苄氨基嘌呤和生长素分别较CK组提高34%、130%和437%。接种GLS后,CK+GLS组玉米素升至0.06 ng/g,水杨酸达峰值,为CK组的18.06倍;脱落酸、6-苄氨基嘌呤和生长素分别为CK组的1.30、2.84和8.78倍;而1-氨基环丙烷-1-羧酸和赤霉素与CK组差异不大。与CK+GLS组相比,LNT+GLS组中6-苄氨基嘌呤、1-氨基环丙烷-1-羧酸、玉米素、赤霉素和生长素分别升高至1.34、1.79、2.99、3.10和3.04倍,而水杨酸和脱落酸分别降至25.24%和8%。聚类分析表明,玉米素和生长素在各处理组中依次升高,聚为一簇;1-氨基环丙烷-1-羧酸和赤霉素在LNT+GLS组中显著积累,聚为另一簇;水杨酸在接种GLS后急剧上升,但LNT处理使其增幅减小,单独成簇。
-
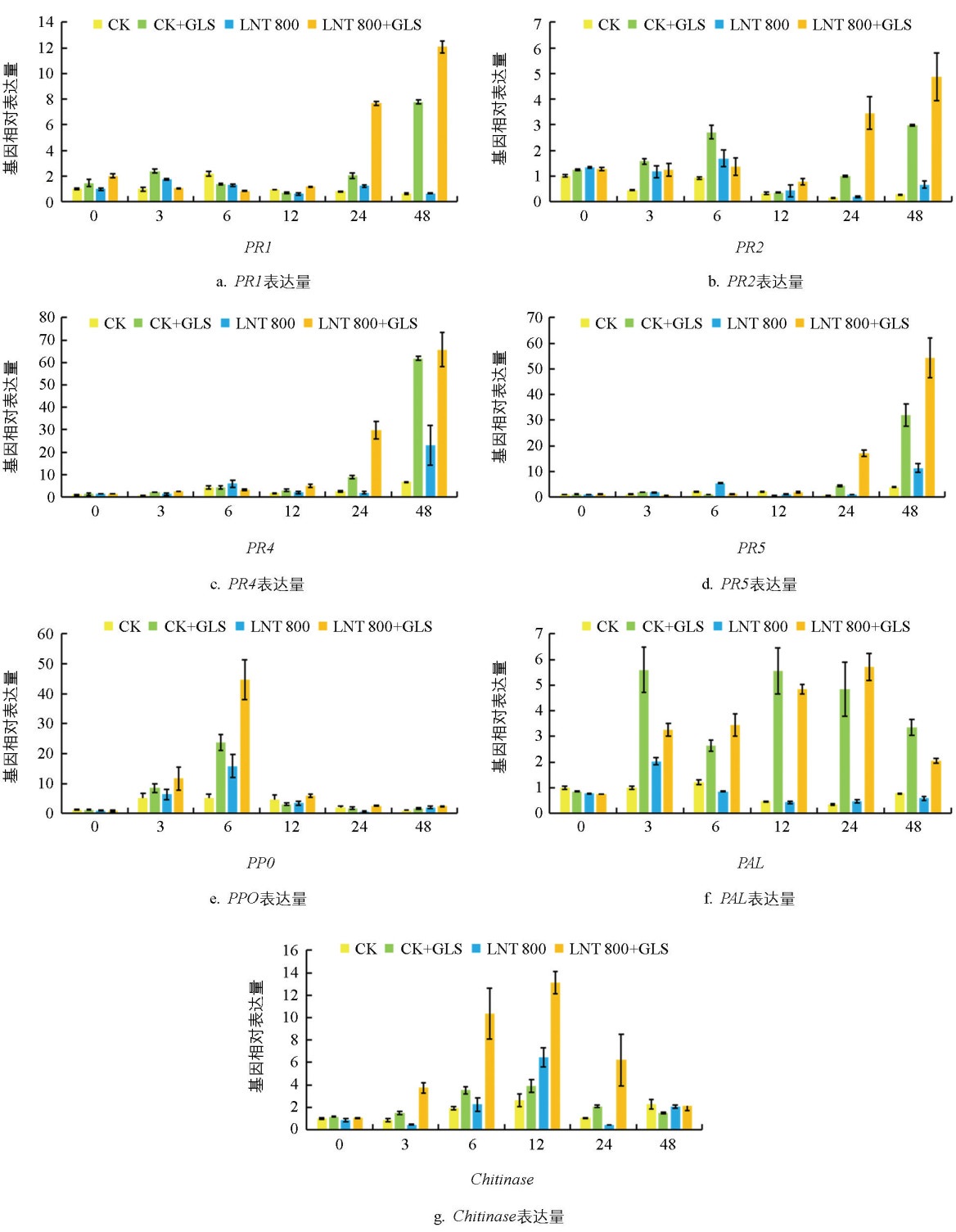
如图 7所示,0 h时各组基因表达无显著差异。接种GLS后,各基因表达均被诱导。PR1基因在24 h和48 h时,LNT+GLS组表达量显著高于其他组,分别为CK组的9.51倍和19.34倍(图 7a)。PR2基因在0~6 h以CK+GLS组表达较高,24~48 h则以LNT+GLS组更高(图 7b)。PR4和 PR5基因表达趋势相似,LNT+GLS组在各时间点均高于CK+GLS组,24 h时分别为后者的3.31倍和3.95倍,48 h时两组均快速上升并达到峰值(图 7 c、d)。PPO基因在3 h和6 h时,LNT+GLS组表达量高于CK+GLS组(图 7e)。PAL基因在3 h时CK+GLS组较高,之后下降,而LNT+GLS组在24 h前缓慢上升(图 7f)。Chitinase基因表达趋势与PPO相似,12 h达峰值(图 7g)。综上,GLS侵染显著诱导7个病程相关基因上调,LNT处理进一步增强其转录水平,提高苹果对GLS的防御能力。
-
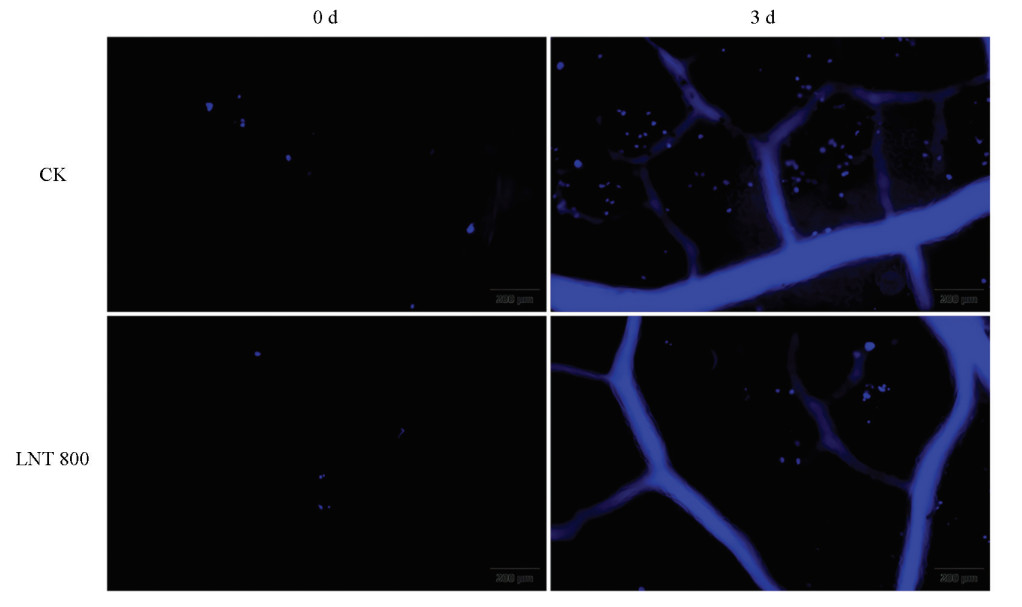
为探究GLS侵染下LNT处理对苹果叶片胼胝质积累的影响,分别对CK+GLS组和LNT+GLS组在第0 d和接种后第3 d的叶片进行胼胝质染色。如图 8所示,接种前(第0 d),两组胼胝质沉积量无显著差异。接种后第3 d,CK+GLS组沉积量显著增强,而LNT+GLS组胼胝质信号强度显著低于同期CK+GLS组。
-
田间喷施800倍LNT稀释液可显著降低苹果叶片病害,病情指数较CK组下降17.96%,免疫诱抗效果达33.56%。同时,株高、新梢数量、新梢长度、东西和南北冠径等生长指标均优于CK组,较CK组分别提高了9.47%、32.79%、28.10%、34.23%和29.52%,其中东西冠径增长最显著(表 2)。表明喷施800倍LNT稀释液可有效降低苹果叶片病害的发病率并促进树体生长。
2.1. LNT对GLS菌丝生长的抑制效果
2.2. LNT喷施处理诱导苹果抗病性的最适浓度筛选
2.3. GLS侵染下LNT对苹果叶片发病情况的影响
2.4. GLS侵染下LNT对苹果叶片显微结构的影响
2.5. GLS侵染下LNT对ROS含量与抗氧化酶活性的影响
2.6. GLS侵染下LNT对苹果叶片中氨基酸、酚类物质和内源激素含量的影响
2.7. GLS侵染下LNT对苹果叶片病程相关基因表达量的影响
2.8. GLS侵染下LNT对苹果叶片胼胝质沉积的影响
2.9. 田间喷施LNT对苹果生长和叶片病情指数的影响
-
平板抑菌试验表明,LNT在200~1 000倍稀释液浓度下均对GLS病原菌的生长表现出一定的抑制作用,但抑制效果未达到显著水平,该结果与已有研究结论一致,即植物免疫诱抗剂主要通过激活系统获得性抗性(SAR)增强植物自身抗病能力,而非直接抑制病原菌生长[19]。离体叶片接种试验进一步表明,喷施800倍LNT稀释液能显著降低苹果叶片GLS发病率和病斑面积,表明该浓度对GLS具有良好防控效果。相对电导率测定结果也证实LNT对叶片细胞膜结构具有保护作用。综上,本研究最终选用800倍LNT稀释液进行后续试验以探究其诱导苹果抗病性的作用机制。
组织学观察发现,CK+GLS组苹果叶片在接种3 d时表皮细胞出现损伤,5 d后结构明显破碎。该损伤源于GLS分生孢子形成附着胞与侵染钉,穿透上表皮并引起细胞壁溶解,同时病原菌分泌酶类降解角质层,破坏表皮防御结构,促进其进一步扩散与定殖。随侵染进展,栅栏组织逐渐褐变,可能与菌丝在胞内扩展有关,最终形成典型褐色坏死斑[20]。相比之下,LNT处理显著延缓了GLS的侵染进程,减轻发病程度,并维持了细胞结构完整性。有报道显示,石莼多糖等植物免疫诱抗剂可通过抑制附着胞黑化阻碍GLS病原菌有效侵染,提示LNT作为多糖类物质可能通过类似机制发挥防控作用[21]。
在植物—病原物互作过程中,ROS暴发是常见的早期响应。为应对氧化胁迫,植物会激活抗氧化酶系统,合成SOD、POD等酶以清除ROS,减轻细胞损伤[22]。本研究显示,GLS侵染诱导苹果叶片中H2O2积累,导致ROS水平升高;而LNT处理增强了SOD和POD活性,显著降低了ROS含量,表明LNT能够激活抗氧化系统,有效缓解由GLS引发的氧化损伤。在激素水平方面,LNT处理显著提高了ABA含量,推测其作为免疫诱抗剂模拟病原菌侵染,激活防御信号、诱导ABA合成和促进气孔关闭,从而限制病原菌侵入[23-24]。另外,LNT处理也使ZT、IAA、ACC和GA3含量有不同程度提高,说明LNT在诱导植物抗病性的同时,可能通过调节生长相关激素促进植物生长发育[25]。ACC作为乙烯前体,其积累可能促使乙烯早期暴发,协同SA信号诱导PR蛋白表达[26]。LNT可能通过提前积累ACC,促使乙烯早期暴发,从而增强对GLS的早期抑制。
氨基酸代谢在植物抗病中发挥重要作用,不仅为蛋白质合成提供基础,还参与次生代谢、能量供应和抗氧化等多种抗病防御反应[27]。本研究发现,LNT处理显著缓解GLS侵染引起的氨基酸消耗,并提高多种抗病相关氨基酸的水平,从而增强苹果对GLS侵染的适应能力。具体表现为:脯氨酸消耗减少,有助于细胞维持渗透平衡和功能稳定[28];精氨酸含量回升,可能通过NO信号途径激活PR基因表达[29];赖氨酸消耗受抑,推测其转化为谷氨酰胺等含氮防御物质参与抗病免疫反应;天冬氨酸含量上升,可能通过三羧酸(TCA)循环增强能量供给[30];甘氨酸作为谷胱甘肽前体含量提高,有助于增强抗氧化能力[31]。综上,LNT通过协同调控氨基酸代谢,在渗透调节、免疫信号、能量代谢和抗氧化防御等多方面增强苹果对GLS的抗性。
酚类物质在植物抗病中既可作为木质素等细胞壁组分的前体增强物理屏障[32],又能通过类黄酮等次生代谢产物直接抑制病原菌生长[33]。本研究显示,LNT处理可促进芦丁、金丝桃苷和槲皮素等类黄酮类物质积累,这些物质可能通过下游代谢增强对GLS的抗性;而CK+GLS组则大量积累阿魏酸和对香豆酸等酚酸类物质,其中阿魏酸参与细胞壁强化[33],对香豆酸作为苯丙烷代谢的关键节点响应病害侵染[34]。因此,LNT可能通过特异性激活类黄酮代谢途径诱导植物系统性抗性,其作用机制不同于病原菌直接侵染引发的广泛酚类积累反应。
在植物免疫中,病原菌侵染会触发病原相关分子模式激发的免疫反应(PTI)与效应蛋白激发的免疫反应(ETI),激活SA等防御信号通路并诱导PR蛋白表达[35]。LNT可能通过模拟病原体相关的分子模式(PAMPs),经模式识别受体(PRRs)激活PTI免疫反应,与病原信号叠加增强防御响应,进而促进SA通路驱动PR基因表达。PR1是植物防御反应的关键标志物[36]。本研究中,GLS侵染诱导SA通路启动并上调PR1表达;LNT+GLS组在12 h后表达量高于CK+GLS组,表明LNT具有免疫诱抗作用。PR2是β-1,3-葡聚糖酶基因,在CK+GLS组早期显著上调,而LNT+GLS组表达较缓,至24 h才明显升高,说明LNT可能延缓了病原体侵染进程。PAL是苯丙氨酸解氨酶基因,在GLS侵染后迅速响应但逐渐减弱,LNT处理则能增强其表达持续性,促进木质素合成以强化细胞壁[37]。
田间试验表明,喷施LNT可显著降低苹果叶部病害病情指数,并有效促进树体生长,表现为东西冠径和新梢数量的显著增加。前人研究也表明LNT具有促进油菜根系生长[38]和提升番茄产量等作用[39],印证了LNT兼具诱导植物抗病性与促进生长的双重功能,符合植物免疫诱抗剂的典型特征。因此,以LNT为核心的植物免疫诱导技术有望成为苹果病害绿色防治的有效策略[40]。然而,LNT的诱抗效果易受品种与田间环境因素影响,且当前研究多限于小区试验,缺乏规模化果园的长期验证,未来需结合区域生态与栽培管理,开展系统性田间集成示范,以推动该技术的产业化应用。
-
本研究采用浓度梯度试验、平板抑菌及离体叶片接种等方法,确定800倍LNT稀释液为外源喷施诱导苹果对GLS抗性的最适浓度。组织学观察表明,LNT处理有助于维持细胞结构完整性,减少防御物质异常沉积。在生理水平上,LNT激活SOD、POD等抗氧化酶活性,缓解氧化损伤;同时调控激素和氨基酸代谢通路,促进黄酮类等抗病次生代谢物积累。基因表达分析表明,LNT诱导多种病程相关基因上调表达。综上,LNT通过协同调控抗氧化防御、激素信号传导和次生代谢等多层次免疫响应,系统增强苹果对GLS的抗性。田间试验进一步证实,喷施800倍LNT稀释液可有效降低病情指数并促进植株生长,展现出良好的应用潜力。