


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
犬弓首蛔虫(Toxocara canis)是一种广泛分布于世界各地的人畜共患寄生线虫,主要寄生于犬和犬科动物的肠道。T. canis生活史复杂,其虫卵随粪便排出后,在适宜的外界环境下发育为感染性虫卵[1]。终末宿主(犬、狐狸、狼等)摄入感染性虫卵后,幼虫经循环系统移行至肠道发育为成虫。人和非特异宿主(鼠、兔、牛等)摄入被感染性虫卵污染的食物、水或含有感染性幼虫的肉制品而感染;幼虫则移行到神经、肌肉、内脏等组织器官,成为滞育的L3幼虫[2-3]。人感染后主要表现为内脏幼虫移行症(Visceral Larva Migrans,VLM)、眼幼虫移行症(Ocular Larva Migrans,OLM)、神经弓首蛔虫病(Neurotoxocariasis,NT)和隐性弓首蛔虫病(Covert Toxocarosis,CT)等多种临床综合征,因此弓首蛔虫病在兽医学及公共卫生学上都应引起足够重视。
原肌球蛋白(Tropomyosin,TM)是一种酸性蛋白,主要由两条α螺旋纤维蛋白链构成[4]。TM存在于多种肌肉组织(如平滑肌、骨骼肌和心肌)中,与肌动蛋白结合形成复合物,参与调节肌肉收缩过程和ATP酶活性,并与肌钙蛋白相互作用构成钙敏感屏障[5-6]。TM具有多种生物学功能,如调节宿主免疫、抑制癌细胞迁移和控制肌肉收缩等。小鼠TM可以促进Th2细胞因子(IL-4、IL-5、IL-10和IL-13)和IgE抗体的释放,从而诱导激活Th2型免疫反应,提高宿主的抗炎能力[7];裸鼠TM4可明显减少胃癌细胞转移瘤数目和肿瘤体积,具有抑制胃癌细胞转移的能力[8]。果蝇(Drosophilidae)TM2与肌钙蛋白复合物结合,可维持肌肉收缩,参与调节肌肉运动[9-10]。
在寄生虫中,TM也参与多种生物学过程,如运动、繁殖、过敏和免疫调节。抑制秀丽隐杆线虫(Caenorhabditis elegans)TM可导致体壁肌肉中的肌动蛋白丝瘫痪和卵巢收缩被抑制,虫体表现为运动缓慢或者瘫痪,以及雌虫无法生殖和产卵等[11]。痒螨(Psoroptes ovis)TM与尘螨过敏患者血清IgE结合,可引发皮肤过敏性炎症反应[12]。小亚璃眼蜱(Hyalomma anatolicum)TM免疫家兔可以引起高滴度的抗体反应[13-15]。目前关于犬弓首蛔虫TM(Tc-TM)的相关研究尚未见报道。
为探讨Tc-TM的免疫保护效果,本试验通过Tc-TM重组蛋白免疫小鼠,检测小鼠体液及细胞免疫应答水平及其抵抗T. canis感染的能力,旨在为寻求T. canis安全高效的亚单位疫苗奠定基础。
全文HTML
-
90只6~7周龄雌性昆明小鼠(体质量约25 g),购自西南医科大学实验动物中心。从动物医院患病犬粪便中收集T. canis成虫,并从雌虫子宫中收集虫卵,在培养皿中培养2~3周,发育至感染性虫卵后收集备用。所有动物饲养及试验方法由西南大学实验动物伦理审查委员会监督(审查号:LAC 2024-2-0170)。
-
重组菌株pET-32a/Tc-TM-BL-21由西南大学分子寄生虫学实验室构建并保存,经IPTG诱导表达和纯化,获得高纯度的Tc-TM重组蛋白后备用。
-
BCA蛋白浓度测定试剂盒、红细胞裂解液和TMB显色液,北京索莱宝科技有限公司;弗氏完全佐剂和弗氏不完全佐剂,碧云天生物技术(上海)有限公司;HRP标记的山羊抗鼠IgG,Sangon Biotech公司;PrimeScriptTM RT reagent Kit、TB Green® Premix Ex TaqTMⅡ和Trizol试剂,Takara公司;CCK-8试剂盒,Biosharp公司。
-
将90只小鼠随机分为6组,即50、75、125 μg/mL Tc-TM免疫组和75 μg/mL pET-32a免疫组、阴性对照组、PBS阳性对照组,每组15只。分别于第0、14、28 d对小鼠进行3次免疫,第42 d时除阴性对照组外每只小鼠经口接种T. canis感染性虫卵(表 1)。于第49 d脱颈法处死小鼠,采集肝脏和肺脏观察病理变化,计算幼虫减虫率,收集各阶段小鼠血清用于后续试验。
-
收集各组小鼠免疫前(0 d)、免疫后(14、28、42 d)及攻虫后第7 d(49 d)的血液样本,离心收集上清。使用2 μg/mL的Tc-TM作为包被液,每孔加入100 μL抗原稀释液置于4 ℃包被过夜。PBST洗涤3次,每孔加入100 μL封闭液;PBST洗涤3次,稀释后的血清样本(1∶ 200)为一抗,山羊抗鼠IgG-HRP(1∶5 000)为二抗,3次PBST洗涤后加入TMB显色液进行显色;使用终止液终止反应,测定OD450 值。
-
在首免后第14 d(14 d)及三免后第14 d(42 d),每组随机选取3只小鼠,无菌分离脾淋巴细胞于RPMI 1640培养液中。PBS阳性对照组使用Con A(10 μg/ mL)刺激。在37 ℃,5% CO2培养箱中培养48 h后提取细胞总RNA,利用qRT-PCR检测细胞因子IFN-γ、IL-2、IL-10、IL-4的mRNA表达水平,每组设置3个重复,引物序列信息见表 2。
-
在攻虫感染后第7 d采集小鼠的肝脏和肺脏,采用人工胃液消化法和改良贝尔曼方法收集幼虫,计算幼虫减虫率(R)。
式中:N阳为PBS阳性对照组平均每鼠检获虫体数;N免为免疫组平均每鼠检获虫体数。
-
在攻虫感染后第7 d采集小鼠的肝脏和肺脏,用4%多聚甲醛固定各组织48 h后,进行脱水、浸蜡、包埋、切片、HE染色并观察各组织的病理变化。
-
采用GraphPad Prism 10.3.1分析各试验数据,结果用x±s表示,两组独立样本之间用t检验,p<0.05为差异有统计学意义。
1.1. 试验动物及虫种
1.2. 菌种及重组蛋白
1.3. 主要试剂
1.4. 试验分组及免疫接种程序
1.5. Tc-TM对小鼠体液免疫水平的影响
1.6. Tc-TM对小鼠细胞因子的影响
1.7. 犬弓首蛔虫幼虫的收集与计数
1.8. 病理组织观察
1.9. 数据分析
-
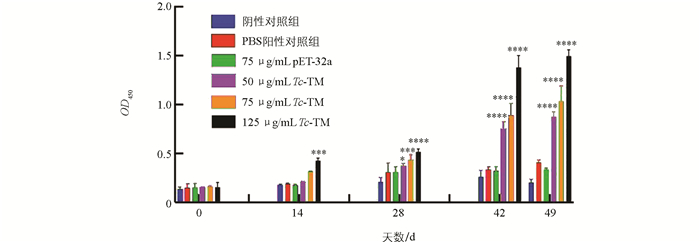
分别用50、75、125 μg/mL Tc-TM免疫小鼠,通过间接ELISA法检测血清中特异性IgG抗体的变化。结果表明,与PBS阳性对照组相比,首免与二免后Tc-TM免疫组IgG抗体含量随免疫次数的增加而上升,三免后Tc-TM免疫组IgG抗体水平达到最高。攻虫后第7 d,各Tc-TM免疫组抗体含量仍维持在较高水平(图 1)。结果表明,Tc-TM能诱导机体产生高滴度的IgG抗体。
-
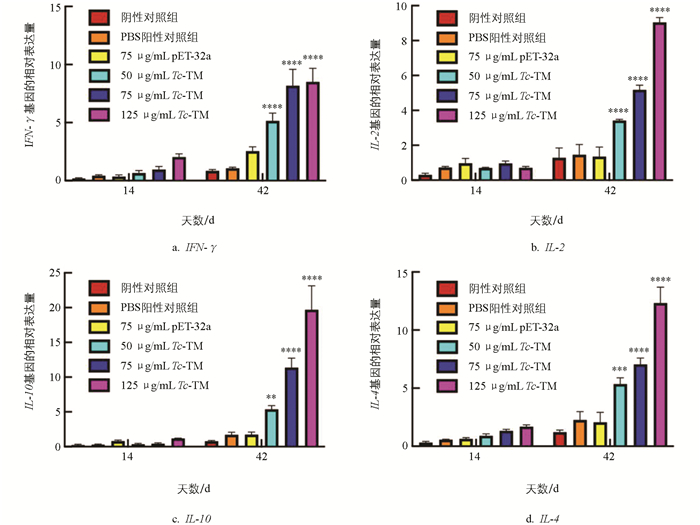
利用qRT-PCR检测细胞因子mRNA的表达水平,结果显示,与PBS阳性对照组相比,首免后第14 d各免疫组细胞因子表达水平均较低(p>0.05)。三免后第14 d,3个免疫组的IFN-γ、IL-2、IL-4和IL-10表达水平均较高(p<0.01)(图 2),且IL-4和IL-10的表达水平显著高于IL-2和IFN-γ(p<0.05),表明免疫后小鼠机体免疫应答趋向于Th2型体液免疫。
-
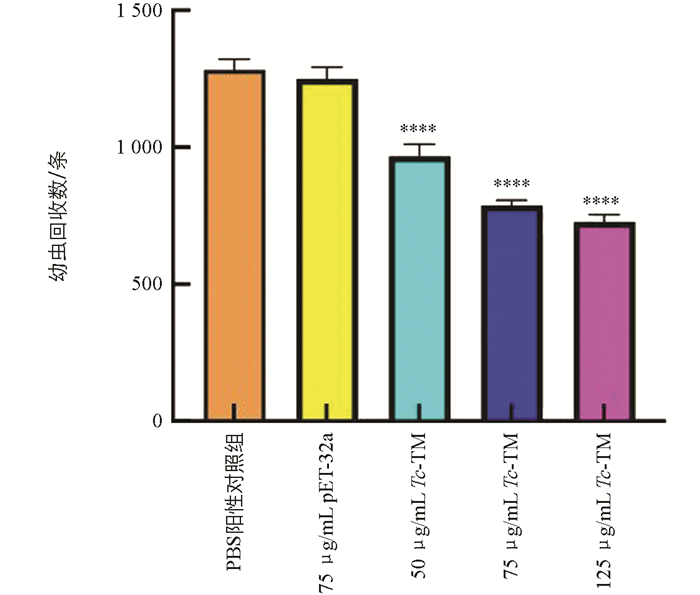
在攻虫后7 d收集肝脏、肺脏的幼虫并计算减虫率,结果显示,与PBS阳性对照组相比,pET-32a免疫组幼虫回收数差异不明显,Tc-TM免疫组肝脏、肺脏的幼虫回收数明显减少,差异有统计学意义(p<0.000 1),减虫率分别为24.6%、38.6%和43.3%(图 3)。
-
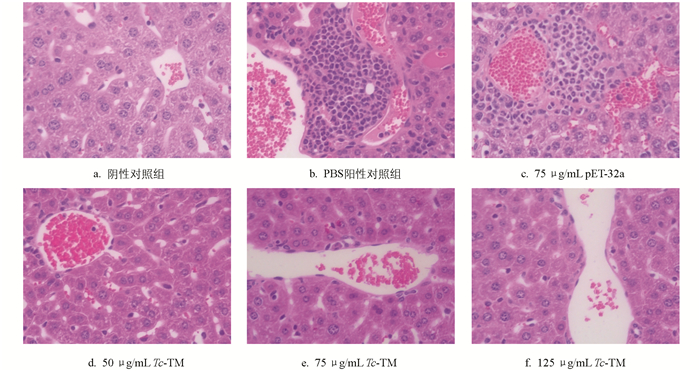
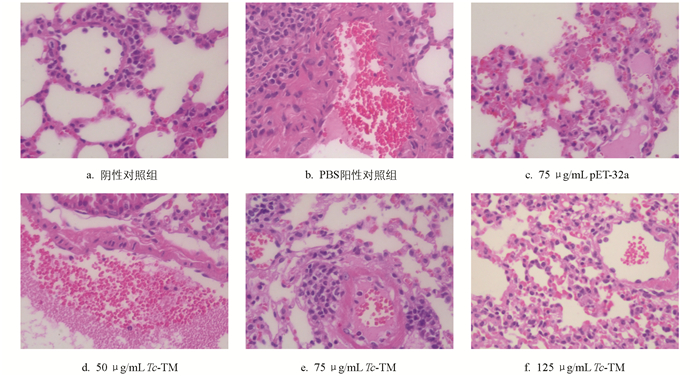
攻虫后第7 d,分离肝脏和肺脏组织制作石蜡切片并进行HE染色观察,结果显示,PBS阳性对照组的肝脏组织中可见大量炎性细胞浸润,主要由淋巴细胞和巨噬细胞组成,呈慢性炎症反应,炎症灶周围的肝细胞出现明显变性,并伴有局灶性坏死(图 4)。肺脏可见大面积炎症细胞聚集和浸润,主要由嗜中性粒细胞和巨噬细胞组成,肺泡轮廓不清(图 5)。Tc-TM免疫组的肝脏和肺脏显示出的炎性浸润和出血点显著减少,炎症聚集程度有所改善,表明重组Tc-TM免疫后可缓解犬弓首蛔虫在宿主体内移行引起的组织损伤。
2.1. 特异性IgG抗体水平的变化
2.2. 细胞因子mRNA的表达水平
2.3. 幼虫减虫率
2.4. 小鼠肝脏和肺脏病理变化
-
TM作为肌动蛋白丝(Actin Filament)的辅助或调控介质,在维持寄生虫结构、介导虫体入侵宿主以及免疫反应中发挥着重要作用。有研究表明,C. elegans TM通过与原肌球调节蛋白相互作用,共同调控细肌丝尖端(Pointed ends)的长度,从而维持肌肉细胞骨架结构的稳定性[14]。疟原虫(Plasmodium falciparum)TM表达降低导致入侵红细胞效率显著下降,严重阻碍P. falciparum的入侵[15]。旋毛虫(Trichinella spiralis)TM能诱导IL-4、IL-5、IL-13、IL-10等Th2型细胞因子的表达,上调IL-10、TGF-β、Arginase-1、CCL2等的表达水平,从而有利于虫体的存活[16]。
IgG是体液免疫中最主要的特异性免疫球蛋白,主要通过细胞毒性(ADCC)和调理吞噬作用在抗寄生虫感染中发挥效应[17]。Hewitson等[18]发现IgG1抗体通过特异性结合多形螺旋线虫(Heligmosomoides polygyrus)分泌的抗原,协同IL-4和IL-25依赖的效应细胞募集髓样细胞包围和限制幼虫迁移发育,从而提供保护性免疫。Venturiello等[19]研究发现IgG包被的T. spiralis幼虫在体外可被嗜酸性粒细胞、中性粒细胞和单核细胞介导的ADCC所杀伤。Clausen等[20]发现P. falciparum子孢子在进入肝细胞前可被循环IgG中和或被补体溶解,从而阻断感染。在本研究中,首免后,IgG抗体水平上升较为缓慢。随着免疫次数的增加,不同质量浓度Tc-TM免疫组的IgG抗体水平显著升高,说明脾脏中相应B细胞克隆的扩增和大量浆细胞已然生成,这正是有效免疫应答的基础。小鼠在T. canis感染后仍维持高滴度的IgG,这可能是Tc-TM诱导了持久的免疫记忆,刺激宿主发动体液免疫应答,从而抵抗T. canis的感染。
寄生虫感染可诱导不同类型的免疫应答,其表现为Th1/Th2型细胞因子的表达水平变化[21]。刚地弓形虫(Toxoplasma gondii)致密颗粒蛋白(GRA7)免疫小鼠后显著上调IFN-γ、IL-2、IL-4的表达水平[22]。日本血吸虫(Schistosoma japonicum)的可溶性虫卵抗原(SEA)免疫小鼠后IL-4、IL-10的表达水平升高,并伴随嗜酸性粒细胞浸润增加,明显诱导Th2型免疫应答[23]。在本试验中,三免后14 d Tc-TM免疫组的Th1型细胞因子IFN-γ、IL-2和Th2型细胞因子IL-4、IL-10转录水平均显著高于PBS阳性对照组(p<0.01),且IL-4和IL-10表达水平显著高于IL-2和IFN-γ(p<0.05),这说明Tc-TM抗原在小鼠体内主要诱导以Th2型为主的Th1/Th2型混合免疫应答。
特异性免疫应答能有效降低寄生虫侵袭宿主组织的能力,从而缓解其引发的病理损伤[24]。Faz-López等[25]研究发现在曼氏血吸虫(Schistosoma mansoni)感染引起的宿主组织损伤修复过程中,Th2型免疫反应通过诱导巨噬细胞分泌抗炎因子与生长因子,显著降低寄生虫的侵袭能力并减少其幼虫数量。另有研究表明,捻转血矛线虫(Haemonchus contortus)TM山羊免疫组与攻虫组相比,成虫数减少48%,虫卵数减少34%,说明TM在抵抗寄生虫感染方面具有显著效果[26]。在本研究中,3个免疫组的幼虫回收数显著减少,减虫率分别为24.6%、38.6%和43.3%,这提示Tc-TM可能诱导高水平的抗体和细胞因子产生,增强了宿主对幼虫的清除作用。HE染色结果显示,Tc-TM缓解了T. canis移行造成的炎性细胞浸润和出血病变,且病变程度随Tc-TM免疫浓度的增加而减轻,说明Tc-TM在小鼠体内发挥了免疫保护作用,有效限制了T.canis的组织侵袭并减轻了宿主的炎症损伤。
-
本研究发现Tc-TM免疫可诱导小鼠产生特异性免疫应答,发挥抗T.canis感染入侵的作用,并减轻了肝脏、肺脏的病理损伤,这一结果为进一步研究犬弓首蛔虫原肌球蛋白的生物学功能奠定了基础。