





-
核因子Y(nuclear factor Y,NF-Y),又称CBF或亚铁血红素激活蛋白HAP,主要由NF-YA,NF-YB及NF-YC 3个亚家族组成[1]. NF-Y蛋白家族的不同成员可通过不同途径进入细胞核,其中NF-YA是由Importin β介导输入核中,而含有HFM亚基的NF-YB和NF-YC作为异二聚体复合物则由Importin 13介导进入细胞核[2]. 3个亚家族可以单独调控基因表达,也可形成NF-YB/NF-YC CONSTANS或NF-Y复合体,进一步结合CCAAT元件参与基因表达的调控[3].
在哺乳动物及酵母体内,NF-Y的3个亚基均由单基因编码[4],但植物中每个亚基均由基因家族编码[5],提供了更多的NF-YA/NF-YB/NF-YC三聚体组合,增加了核因子Y功能的复杂性,参与了在种子萌发、胚胎发育、开花过程以及植物对氮、磷、脱落酸(ABA)以及盐分胁迫等的响应过程[6]. 研究表明大豆GmNF-YCa在蔗糖和甘露醇诱导后上调表达,过表达可显著改善甘露醇和蔗糖胁迫下拟南芥种子的发芽率、根长和侧根伸长明显等[7]. 谷子SiNF-YA6主要在根部表达,同时受低氮、干旱、高盐和ABA等胁迫的诱导表达,该基因的过表达显著提高了拟南芥对低氮胁迫的耐受性[8]. 由此可见,NF-Y蛋白家族的不同成员不仅参与了植物多种生长发育进程,且在植物抵御逆境胁迫等方面具有重要作用.
太子参为传统中药,系石竹科植物孩儿参Pseudostellaria heterophylla (Miq.) Paxex Pax et Hoffm的干燥块根,含有多糖类、皂苷类、磷脂类、环肽类等活性成分,具有增强免疫力、降血糖、抗疲劳等多种功效[9]. 随着近年来太子参需求量的逐年上升,逐步转向人工种植,主要分布于温暖湿润的气候环境中[10]. 高温、干旱等逆境容易引起太子参生长发育受阻,从而导致减产、病害等问题. 因此,研究NF-Y基因家族的表达模式及对干旱胁迫的响应,对于深入理解其在太子参生长发育过程中的功能和作用,解析NF-Y蛋白在植物抵抗逆境胁迫中的作用机制具有重要意义.
HTML
-
从NCBI中下载甜菜(Beta vulgaris L.)、拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa L.)等的NF-Y蛋白家族成员. 通过BlastP分别比对3个数据库,从中鉴定太子参NF-Y家族同源序列,期望值设为小于0.001,结合数据库原有的注释结果,剔除注释错误的序列,将3个数据库中鉴定的NF-Y蛋白序列统一编号.
-
采用muscle 3.8软件(http://www.drive5.com/muscle/)对太子参及甜菜、拟南芥及水稻进行多序列比对,分析结果用MEGA X(https://www.megasoftware.net/)分析,采用邻接法(neighbor-joining method)构建NF-Y的系统进化树,检验参数Booistrap重复1 000次. 利用MEME在线程序(http://meme-suite.org/)对NF-Y蛋白家族的保守基序进行分析,预测太子参NF-Y蛋白家族的蛋白质保守基序(Motif),参数设置为默认值,并用WebLogo软件(http://weblogo.berkeley.edu/logo.cgi)展示保守基序.
-
采用RSEM软件(http://deweylab.biostat.wisc.edu/rsem/)进行基因表达分析,采用RPKM值(reads per kb permillion reads)表示基因的相对表达水平,采用Cluster 3.0软件对NF-Y基因的表达水平进行标准化,最后采用Heatmap图展示基因的表达.
-
采用美吉生物云平台(https://cloud.majorbio.com/)中的STRING数据库(http://string-db.org/)对NF-Y蛋白互作进行分析. 参考拟南芥的蛋白互作关系,构建太子参NF-Y蛋白的互作关系网络. 采用confidence score值评价互作关系的可信度,默认值0.4,值越大代表互作关系越可靠.
2.1. NF-Y基因家族的鉴定
2.2. 系统进化树的构建及蛋白保守序列比对
2.3. 表达分析
2.4. 蛋白互作分析
-
采用拟南芥、水稻等模式植物的NF-Y蛋白检索3个太子参转录组数据库,结合注释信息及手工分析,共鉴定出24个NF-Y基因(表 1),分别归入3个亚家族,包括9个NF-YA,10个NF-YB及5个NF-YC. 其中数据库1中获得9个NF-YA,6个NF-YB及4个NF-YC;数据库2中获得8个NF-YA,8个NF-YB及4个NF-YC;数据库3中获得8个NF-YA,7个NF-YB及4个NF-YC. PhNF-YA8仅在数据库1中检索到,PhNF-YB5,PhNF-YB6,PhNF-YB7,PhNF-YB9及PhNF-YC5未在数据库1中检索到,暗示这些基因可能是诱导型基因.
-
为了推测24个太子参NF-Y蛋白的功能,分别构建了NF-YA,NF-YB及NF-YC氨基酸序列的系统进化树(图 1). 太子参的9个NF-YA蛋白分别归入3个类群,其中PhNF-YA2,PhNF-YA3,PhNF-YA7,PhNF-YA9与AtNF-YA3,AtNF-YA5,AtNF-YA6,AtNF-YA8归为第1类群,PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA8与AtNF-YA1,AtNF-YA9归为第2类群,PhNF-YA6与AtNF-YA2,AtNF-YA10归入第3类群.
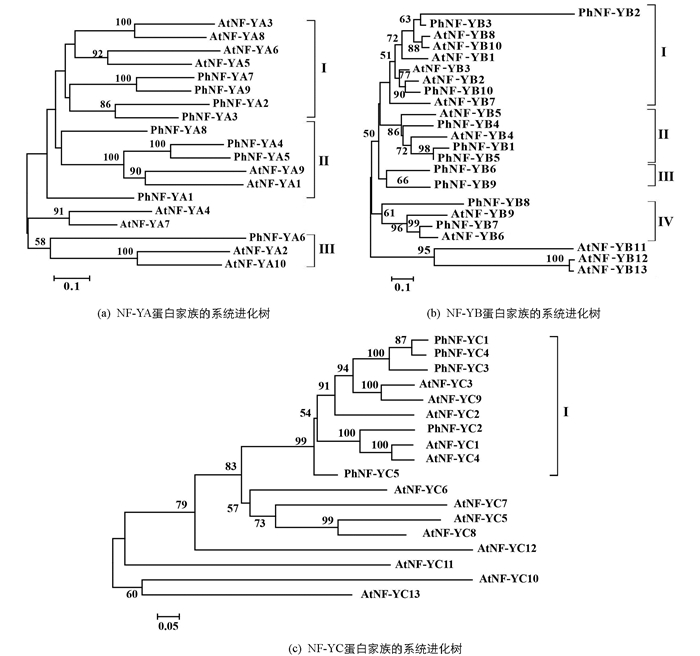
太子参的10个NF-YB蛋白分别归入4个类群,其中PhNF-YB2,PhNF-YB3,PhNF-YB10与AtNF-YB1,AtNF-YB2等6个拟南芥NF-YB蛋白归入第1类群,PhNF-YB1,PhNF-YB4,PhNF-YB5与AtNF-YB4,AtNF-YB5归入第2类群,PhNF-YB6及PhNF-YB9归入第3类群,PhNF-YB7,PhNF-YB8与AtNF-YB6,AtNF-YB9归入第4类群. 太子参的5个PhNF-YC蛋白归入同一个类群.
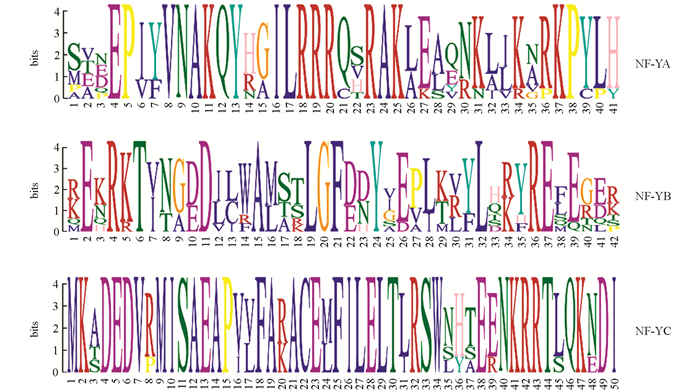
为了解太子参NF-Y蛋白的序列特点,分析了太子参NF-Y蛋白的保守基序(图 2),发现PhNF-YA,PhNF-YB及PhNF-YC亚家族中均具有一段高度保守的序列,暗示这些序列与太子参NF-Y蛋白的功能密切相关.
-
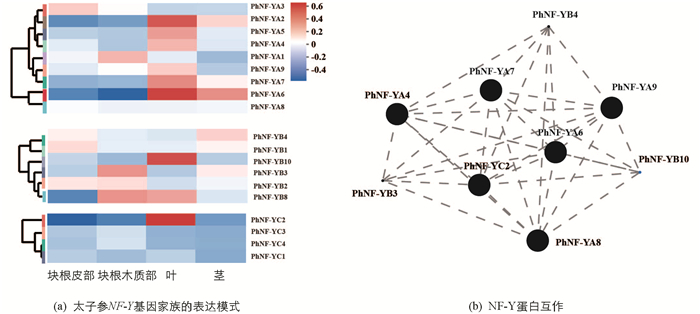
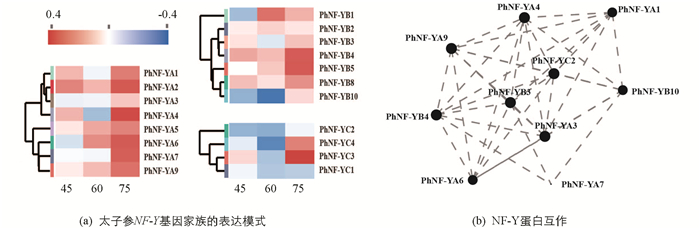
为了进一步阐明太子参NF-Y蛋白的功能,分析了这些基因在叶、茎、块根皮部及块根木质部中的表达模式(图 3a),结果显示9个PhNF-YA基因中PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9仅在叶中有较高表达,PhNF-YA2,PhNF-YA6在叶及茎中均有较高表达,PhNF-YA1在块根木质部有较高表达,PhNF-YA3在块根皮部有较高表达. 6个PhNF-YB基因中PhNF-YB1和PhNF-YB4在块根皮部及茎中有较高表达,PhNF-YB10仅在叶中有较高表达,PhNF-YB3在块根木质部中有较高表达,PhNF-YB8在块根木质部及叶中有较高表达. 4个PhNF-YC基因中PhNF-YC2在叶中有较高表达,其余基因表达量较低. 19个基因的蛋白互作关系分析结果显示(图 3b),PhNF-YA4及PhNF-YC2等9个PhNF-Y蛋白的互作关系在太子参叶片的生长发育过程中具有重要作用.
-
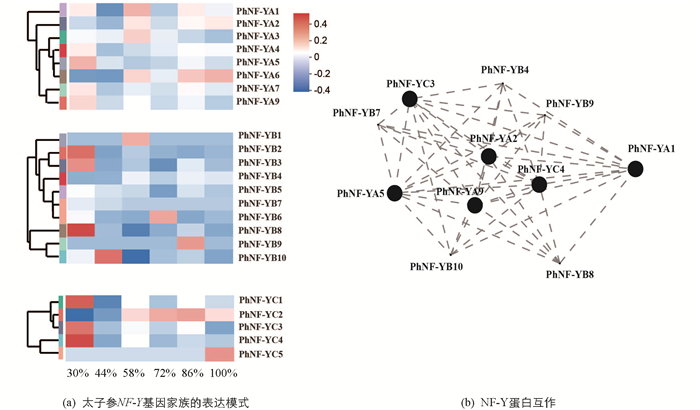
为了研究太子参NF-Y蛋白对水分胁迫的响应机制,分析了不同土壤水分胁迫下块根中NF-Y基因的表达模式(图 4a),结果显示8个PhNF-YA基因中PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9在干旱胁迫下(30%)有较高表达,PhNF-YA1,PhNF-YA2,PhNF-YA3在低度干旱条件下(58%)有较高表达,PhNF-YA6随着土壤水分含量的上升表现出上调的趋势. 10个PhNF-YB基因中PhNF-YB2,PhNF-YB3及PhNF-YB8在干旱胁迫下(30%)有较高表达,PhNF-YB10在适度干旱胁迫下(44%)有较高表达,PhNF-YB1在低度干旱条件下(58%)有较高表达. 5个PhNF-YC基因中PhNF-YC1,PhNF-YC3,PhNF-YC4在干旱胁迫下(30%)有较高表达,PhNF-YC2与土壤水分的增加趋势基本一致,PhNF-YC5仅在涝滞环境下(100%)有较高表达. 23个PhNF-Y蛋白的互作关系表明(图 4b),PhNF-YA1,PhNF-YB4,PhNF-YC3及PhNF-YC4等11个PhNF-Y蛋白积极响应土壤水分的胁迫,调控植物的生长发育.
-
为了研究太子参NF-Y蛋白对蔗糖诱导的响应机制,利用组培苗观察不同蔗糖浓度培养条件下太子参根部NF-Y蛋白的表达模式(图 5a),结果显示,随着培养基中蔗糖浓度的升高,3个PhNF-YA基因的表达呈现逐渐升高的趋势,7个PhNF-YB基因中PhNF-YB5的表达量随蔗糖浓度的升高而升高,PhNF-YB1,PhNF-YB2的表达呈先上升后下降的趋势,PhNF-YB3,PhNF-YB4,PhNF-YB8,PhNF-YB10的表达呈先下降后上升的趋势. 4个PhNF-YC基因中PhNF-YC3及PhNF-YC4基因的表达受高浓度蔗糖诱导,PhNF-YC1在高浓度蔗糖条件下表达受到抑制. 19个PhNF-Y蛋白的互作关系表明(图 5b),PhNF-YA1,PhNF-YB4,PhNF-YC2等10个PhNF-Y蛋白积极响应蔗糖胁迫,协同调控植物的生长发育.
3.1. 太子参NF-Y家族基因的鉴定与生物信息学分析
3.2. 太子参NF-Y家族基因蛋白的保守基序及进化分析
3.3. 不同组织中NF-Y家族基因的表达分析
3.4. 块根中NF-Y家族基因对土壤水分胁迫的响应
3.5. 根中NF-Y家族基因对蔗糖胁迫的响应
-
已有证据表明NF-Y蛋白在叶绿体形成及光合作用中具有重要作用. 拟南芥AtNF-YA5,AtNF-YB9和AtNF-YC9形成的复合体可与GCR1,GPA1和Pirin1组成信号传递链,参与调控植株中叶绿素a/b结合蛋白基因的表达[14]. 过量表达小麦TaNF-YB3后显著提高叶绿素含量、光合速率及早期生长率[15]. 水稻Os-NF-YB2/3/4可影响多个核酮糖-1,5-二磷酸羧化酶/氧合酶小亚基和叶绿素a/b结合蛋白基因的表达[16]. 本研究发现太子参PhNF-YA4,PhNF-YA5等6个基因,PhNF-YB8,PhNF-YB10及PhNF-YC2在叶中高量表达,且可能具有复杂的相互作用关系,暗示这些基因在协同调控叶的生长发育及光合作用中具有重要作用,但具体机制尚不明确. 拟南芥NF-YA1/5/6/9共同调控雄配子发育、胚胎发育、种子形态及萌发等[17-18],单个基因或两个基因的突变并不能发生表型的改变,但过表达其中任一基因均能影响种子的发育,暗示了太子参PhNF-YA亚家族也有功能冗余的可能性. 此外,PhNF-YA1及PhNF-YB3,PhNF-YB8在块根木质部中有较高表达,且进化分析结果显示PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA8与AtNF-YA1,AtNF-YA9聚为一类,暗示了这些太子参NF-Y基因可能与太子参块根的发育有一定关系.
NF-Y蛋白在水分胁迫过程中具有重要作用. 研究表明拟南芥大部分NF-YA基因呈上调表达,而小麦中NF-YA则相反[19-20]. 过表达AtNF-YB1和ZmNF-YB2可显著减少转基因植株的叶片水分流失及提高抗旱能力[21]. 部分NF-Y基因的转录受ABA途径调控,转录后受到miR169的调控[22]. 本研究中太子参PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9,PhNF-YB2,PhNF-YB3,PhNF-YB8及PhNF-YC1,PhNF-YC3,PhNF-YC4在干旱胁迫下均显著上调,且进化分析结果显示PhNF-YB2,PhNF-YB3,PhNF-YB10与AtNF-YB1,AtNF-YB2等6个拟南芥NF-YB蛋白归入一类,暗示了这些太子参NF-YB蛋白与抗旱有密切联系. 此外,蛋白间互作关系暗示PhNF-YC3,PhNF-YC4在干旱胁迫响应过程中可能具有核心调控作用. 值得注意的是,PhNF-YA6及PhNF-YC2的表达随土壤水分的增加而增加,揭示了这些基因在太子参感知土壤水分变化的过程中具有重要作用.
蔗糖及其衍生物不仅是光合同化物与能量的运输和贮藏形式,也是植物中一种重要的信号分子,调控基因表达及影响器官的生长发育[23-24]. 本研究分析了太子参NF-Y蛋白对蔗糖胁迫的响应,发现了大部分基因能够响应蔗糖浓度的变化,暗示了环境因子可通过影响组织中蔗糖的分布来调控NF-Y蛋白的表达,进而影响器官的形态建成.
 DownLoad:
DownLoad: