





-
开放科学(资源服务)标志码(OSID):

-
李(Prunus salicina Lindl.)为蔷薇科李属植物,是我国南北方皆宜种植的温带落叶果树,近年来,作为重庆市三大水果之一的脆李种植面积和产量连续增长,截至2021年6月,全市李种植面积达到9.59万公顷,成为继柑橘之后的第二大水果,其中以巫山脆李为代表的青脆李,具有果形端庄、肉质脆嫩、味甜多汁等特点,约占全市李种植面积的80%,深受消费者的喜爱. 目前,重庆青脆李苗木繁殖用砧木以毛桃为主,生产上存在易感根瘤、天牛危害严重、易流胶、易早衰、耐涝性差等问题[1]. 因此,筛选替代性李砧木,对于推动青脆李产业可持续发展具有十分重要的意义. 研究小组近年对重庆地区野生李分布情况进行了调查研究,发现巫溪、城口、南川等地分布有大量的野生李,生长于溪流灌丛边或林边,海拔700~1 800 m,其枝条上多有退化的针刺,果实小,味酸,果皮苦涩而不堪食用. 在巫山、巫溪等脆李主产区,有果农自发挖取大山中的野生李根蘖苗或实生小苗做砧木嫁接青脆李,比毛桃砧木的树体寿命更长,病虫害相对较轻. 本研究以巫山脆李的一年生枝条为接穗,嫁接在4种不同砧木上,通过嫁接亲和性评价、接穗生长量测量、嫁接口切片观察和嫁接亲和性相关基因表达量分析,综合评价野生李作为巫山脆李砧木的可行性,以期为后续巫山脆李砧木育种提供重要理论依据.
HTML
-
巫山脆李接穗由巫山县果品产业发展中心提供. 毛樱桃(Cerasus tomentosa Thunb. Wall)、毛桃(Amygdalus persica L. Batsch)和山杏(Armeniaca sibirica L. Lam)一年生砧木苗由北京市农林科学院林业果树研究所提供,野生李(Prunus salicina Lindl.)一年生砧木为研究团队收集的巫溪野生李种子自行繁育. 砧木苗于2020年冬季用营养钵栽植,在盆底部放入少量碎石后用腐殖质基质栽培,盆深20 cm,上底直径21.3 cm,下底直径18 cm,底部开有漏水孔,2021年春季(2月初)进行嫁接试验.
-
在嫁接后第43 d、50 d进行测定,其后每隔10 d测定一次,共测定7次. 接穗长度:采用卷尺进行测量,测量嫁接口到接穗顶端的长度. 接穗粗度:采用游标卡尺测量嫁接口接穗上5 cm处的枝梢直径. 嫁接成活率在嫁接后第50 d统计.
-
在嫁接后50 d时,采取从顶端往下数第6片无病虫害、叶色正常、新抽发并转绿春梢叶片,用直尺测量功能叶片的叶长、叶宽,用电子天平称量新鲜叶片的鲜质量,然后置于烘箱中105 ℃杀青15 min,调温至70℃烘干至恒重并称取叶片干质量. 参照聂佩显等[2]的方法,测定比叶鲜质量、比叶干质量.
-
取嫁接后50 d的植株叶片各3片用于制作石蜡切片,先用流水将叶片表面杂质冲洗干净,用纸巾吸干水分或自然风干,将叶片中部横向剪切下来,完全浸入FAA固定液(70%无水乙醇∶冰乙酸∶福尔马林=18∶1∶1)中,期间每隔24 h换一次固定液,直至固定液不变色,叶片脱色至无色为止. 参照李和平[3]的方法进行石蜡切片观察.
-
在嫁接后50 d左右,砧穗间维管组织建立连接,此时砧穗紧密粘合在一起,采集砧穗间愈伤组织以及嫁接口茎段,参照孟纯阳[4]的方法硬组织包埋和切片,在显微镜下拍照,以观察此时不同砧穗间愈合情况.
-
光合特性测定采用ADC LC 20pro光合仪测定. 于嫁接后第50 d,选取每个植株4个方位从顶端往下数第6~8片功能叶(无病害、叶色正常、叶片无缺损)为试验材料,参考张涵[5]的方法测定净光合速率、气孔导度、胞间CO2浓度.
-
在光合参数测定结束后,采摘所测定的功能叶片,并做好标记带回实验室. 先用流水洗净叶片上下表面的杂质,用纸巾吸干水分. 参照张宪政[6]、叶济宇[7]的方法测定叶绿素a、叶绿素b、类胡萝卜素、总叶绿素、总光合色素等.
-
小心刮取嫁接口愈伤组织,参照Beauchmp等[8]和Zhou[9]等的方法测定SOD活性,参照Muñoz-Muñoz等[10]和Quintanilla-Guerrero等[11]的方法测定POD活性,参考Sinha等[12]的方法测定CAT活性,参考李合生[13]的方法测定可溶性蛋白含量.
-
取嫁接后第50 d砧穗间的愈伤组织,进行砧穗间亲和性相关基因qRT-PCR分析. RNA的提取采用TIANGEN RNA prep Pure Plant Plus Kit试剂盒,引物由北京擎科生物科技有限公司合成(表 1),使用德国耶拿Analytik-jena qTOWER3 G qPCR仪进行基因表达水平分析,循环参数为94 ℃ 1 min; 94 ℃ 10 s,60 ℃ 30 s,72 ℃ 1 min,40个循环; 最后反应产物进行熔解曲线分析(60 ~95 ℃),鉴定产物特异性,根据Ct值计算基因的相对表达水平. 以李Actin基因为内参基因,每一个基因设置3次重复. 延伸过程中收集荧光信号,最后进行溶解曲线的绘制,并计算各个反应的RQ值.
1.1. 试验材料
1.2. 植株生长量和嫁接成活率测定
1.3. 叶片形态及比叶重比较
1.4. 叶片解剖结构观察
1.5. 嫁接部位愈合情况观察
1.6. 叶片光和参数测定
1.7. 叶片光合色素含量测定
1.8. 嫁接口亲和性相关酶活性测定
1.9. 嫁接口愈伤组织亲和性相关基因荧光定量qRT-PCR检测
-
嫁接后每10 d手工剥离嫁接口观察,在嫁接后50 d,发现嫁接口愈伤组织生长良好,砧木与接穗贴合较好,故选取该时间统计4种砧木的嫁接成活率. 由表 2可以看出,野生李砧木嫁接成活率最高,达到99.03%; 其次是毛桃和山杏砧木,成活率分别是88.64%和70.00%,山樱桃作砧木的嫁接成活率最低,仅为46.94%. 从嫁接成活率上来看,野生李和毛桃更适合作巫山脆李的砧木,山杏和山樱桃的嫁接成活率较低,不适宜作巫山脆李的砧木.
-
通过硬组织包埋技术,对嫁接口茎段进行纵向切片,观察嫁接后50 d山樱桃、毛桃、野生李和山杏4种砧木与接穗的嫁接口愈合情况,结果如图 1所示. 从图中可以看出,山樱桃(图 1a)砧木嫁接苗砧穗间薄壁细胞细胞壁木质化明显; 毛桃(图 1b)和野生李(图 1c)两种砧木嫁接处出现了大量的薄壁细胞,未观察到明显的木质化现象,砧木与接穗愈合良好; 山杏(图 1d)砧木嫁接后,砧木与接穗之间细胞出现黑化现象,初步判断是薄壁细胞凋亡,有少量的细胞壁木质化现象,表明砧木与接穗之间部分愈合不良. 由此可见,巫山脆李与野生李、毛桃之间愈合良好,与山杏和山樱桃之间愈合不良.

-
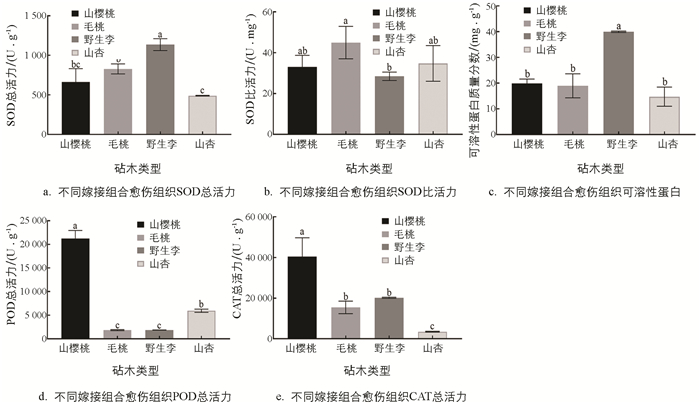
巫山脆李不同砧木嫁接组合砧穗间愈伤组织不同酶活性如图 2所示. 野生李砧木嫁接口愈伤组织的超氧化物歧化酶(SOD)总活性力高,其次是毛桃砧木,最低的是山杏砧木(图 2a). 毛桃砧木嫁接口愈伤组织的SOD比活力最高,其次是山樱桃和山杏砧木,野生李砧木最低(图 2b). 野生李砧木嫁接口愈伤组织的可溶性蛋白质量分数显著高于其余3种砧木,其余3种砧木愈伤组织种可溶性蛋白质量分数差异不具有统计学意义(图 2c). 山樱桃砧木嫁接口愈伤组织的过氧化物酶(POD)总活力最高,其次是山杏砧木,野生李和毛桃最低(图 2d). 山樱桃砧木嫁接口愈伤组织的过氧化氢酶(CAT)总活力最高,其次是毛桃和野生李砧木,且两者之间CAT总活力没有差异,山杏砧木最低(图 2e).
-
对不同砧木嫁接组合砧穗间愈伤组织亲和性相关基因进行定量分析,结果表明,毛桃砧木嫁接苗砧穗间愈伤组织生成关键调控基因WIND1 (图 3a)相对表达量最高,其次是野生李砧木,山樱桃和山杏砧木嫁接苗砧穗间愈伤组织相对表达量最低,毛桃砧木嫁接苗砧穗间愈伤组织WIND1 基因相对表达量是野生李砧木嫁接苗的1.35倍,是山樱桃砧木嫁接苗的2.56倍,是山杏砧木嫁接苗的2.99倍. 野生李砧木嫁接苗砧穗间愈伤组织的WIND1 基因表达量是山樱桃砧木嫁接苗的1.89倍,是山杏砧木嫁接苗的2.21倍,山樱桃和山杏砧木嫁接苗砧穗间愈伤组织的WIND1 基因相对表达量差异不具有统计学意义. 山樱桃和山杏砧木嫁接苗砧穗间愈伤组织苯丙氨酸解氨酶基因PAL1 相对表达量(图 3b)最高,显著高于毛桃和野生李砧木嫁接苗,毛桃和野生李砧木嫁接苗砧穗间愈伤组织PAL1 基因相对表达量差异不具有统计学意义. 野生李砧木嫁接苗砧穗间愈伤组织的转录因子ARF1 相对表达量(图 3c)最高,其次是山樱桃和山杏砧木嫁接苗,毛桃砧木嫁接苗砧穗间愈伤组织的转录因子ARF1 相对表达量最低. 野生李砧木嫁接苗砧穗间愈伤组织木质素生成途径关键酶基因 4CL1、4CL2和4CL3 相对表达量(图 3d)最高,其次是山樱桃和毛桃砧木嫁接苗,山杏砧木嫁接苗砧穗间愈伤组织 4CL1 基因相对表达量最低. 综上所述,由亲和性基因定量结果可以看出,与巫山脆李亲和性最强的是野生李和毛桃,山杏和山樱桃与巫山脆李之间亲和性较差.

-
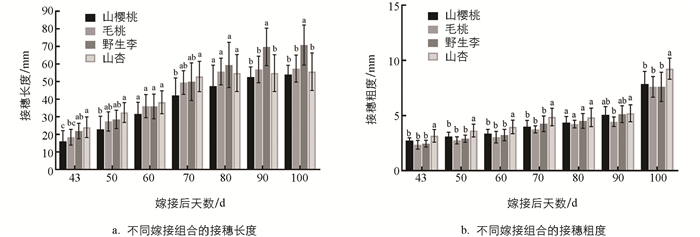
从嫁接后43 d开始到嫁接后100 d,进行了7次接穗长度和粗度测量,结果如图 4所示. 从图中可以看出,同一品种接穗在不同砧木上的生长量存在明显差异. 山樱桃和毛桃作砧木的嫁接苗生长量较小,枝梢长度和粗度均显著低于野生李和山杏砧木嫁接苗,具有植株矮、茎干细的特点; 野生李作砧木的嫁接苗,接穗生长最快,具有植株高、茎干细的特点; 山杏作砧木的嫁接苗,枝梢最粗,具有植株矮、茎干粗的特点. 综上所述,从植株早期生长情况来看,野生李和毛桃砧木均适合作巫山脆李的砧木,嫁接苗的生长势均较好.
-
4种砧木嫁接苗的叶片外观形态如图 5所示,不同砧木的嫁接苗叶片形态指标有差异(表 3),山樱桃作为砧木的嫁接苗叶片呈长椭圆形(图 5a),毛桃(图 5b)、野生李(图 5c)和山杏(图 5d)作为砧木的嫁接苗叶片均呈倒卵形,其中,野生李作砧木嫁接苗的叶片长度和宽度明显大于毛桃和山杏作砧木的嫁接苗.

对4种不同砧木嫁接苗功能叶的比叶鲜质量和比叶干质量测定结果如图 6所示. 从图中可以看出,山樱桃比叶鲜质量(图 6a)和比叶干质量(图 6b)均显著高于其余3种砧木的嫁接苗,毛桃、野生李和山杏作砧木的3种嫁接苗比叶鲜质量和比叶干质量均不具有统计学意义. 由此判断,山樱桃砧木嫁接苗叶片厚度和密度要大于毛桃、野生李和山杏砧木嫁接苗.

-
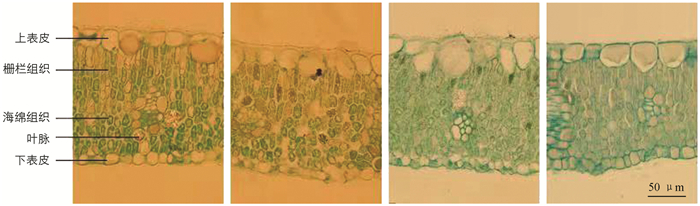
对叶片石蜡切片的显微观察结果如图 7所示,山樱桃(图 7a)砧木的嫁接苗叶片有3~4层栅栏组织细胞,毛桃(图 7b)砧木的嫁接苗叶片的栅栏组织只有2层,野生李(图 7c)砧木嫁接苗叶片的栅栏组织有2~3层,山杏(图 7d)砧木嫁接苗叶片的栅栏组织有3层. 由图 8可见,栅栏组织厚度方面,山杏砧木嫁接苗最厚,其次是野生李和山樱桃砧木嫁接苗,毛桃砧木嫁接苗叶片栅栏组织最薄(图 8c). 毛桃砧木嫁接苗叶片、栅栏组织厚度与海绵组织厚度对比(图 8f)、叶片组织结构紧密度(图 8g)、栅栏组织厚度占叶肉组织的比例(图 8i)均低于其余3种砧木嫁接苗叶片,海绵组织厚度(图 8d)、叶片组织结构疏松度(图 8h)高于其余3种砧木嫁接苗叶片,上表皮厚度(图 8a)与其余3种砧木嫁接苗叶片没有差异. 综上所述,毛桃砧木嫁接苗的叶片具有叶片薄、栅栏组织少、海绵组织多、叶片结构疏松的特点; 山杏砧木嫁接苗具有下表皮厚度、海绵组织厚度、叶片厚度、叶片组织结构紧密等特点. 前人的研究结果表明,植物的叶片栅栏组织厚度和叶片组织结构紧密度与植物耐旱性成正比,可以看出山杏砧木嫁接苗对干旱的耐受度可能要比其他3种砧木嫁接苗强.

-
在固定光强1 000 μmol/m2·s下,不同砧木嫁接苗在嫁接后第50 d测定的光合参数数据如图 9所示. 毛桃砧木嫁接苗净光合速率最高,为11.75 μmol/m2·s; 其次是山樱桃和山杏砧木嫁接苗,野生李砧木嫁接苗净光合速率最低,为3.20 μmol/m2·s(图 9a). 气孔导度方面,毛桃砧木嫁接苗气孔导度最高,为0.16 mmol/m2·s,其次是山樱桃砧木嫁接苗,野生李和山杏砧木嫁接苗最低(图 9b). 胞间CO2浓度方面,山樱桃和野生李砧木嫁接苗胞间CO2浓度最高,其次是毛桃砧木嫁接苗,山杏砧木嫁接苗胞间CO2浓度最低(图 9c).

-
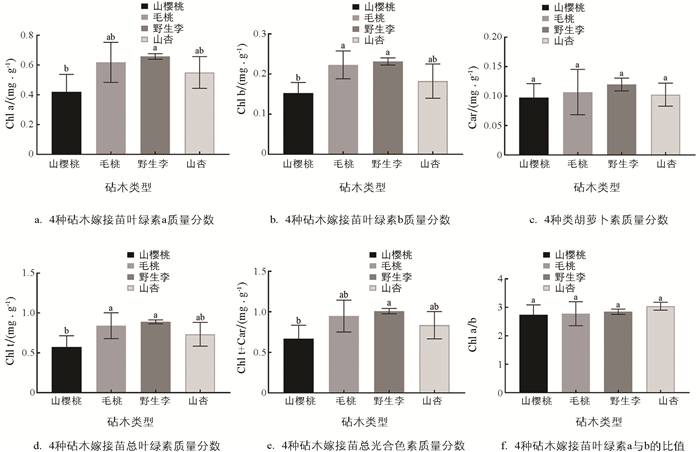
植物光合作用主要参与色素有叶绿素a、b(Chl a、b)和类胡萝卜素(Car). 4种砧木嫁接苗在嫁接后第50 d测定的叶片光合色素的质量分数如图 10所示,从图中可以看出,单位质量Chl a、Chl b、Car、总叶绿素(Chl t)和总光合色素(Chl t+Car)含量以野生李砧木嫁接苗最高,山樱桃嫁接苗最低,毛桃嫁接苗含量略低于野生李嫁接苗,但差异不具有统计学意义(图 10a-e). 4种砧木嫁接苗的Chl a/b比值差异不具有统计学意义(图 10f). 综上所述,野生李和毛桃砧木嫁接苗叶片光合色素质量分数高于山杏和山樱桃砧木嫁接苗,这可能是毛桃和野生李砧木嫁接苗生长量和光合速率高的原因之一.
2.1. 4种砧木与巫山脆李的亲和性评价
2.1.1. 嫁接成活率对比
2.1.2. 嫁接口细胞学观察
2.1.3. 砧穗间愈伤组织亲和性相关酶活性测定
2.1.4. 砧穗间愈伤组织亲和性相关基因qRT-PCR分析
2.2. 4种砧木嫁接巫山脆李的生长表现
2.2.1. 不同砧木嫁接苗早期生长情况
2.2.2. 不同砧木嫁接苗叶片形态及比叶重比较
2.2.3. 不同砧木嫁接苗叶片解剖结构观察
2.3. 4种砧木嫁接苗的光合生理指标比较
2.3.1. 不同砧木嫁接苗植株光合特性分析
2.3.2. 不同砧木嫁接苗叶片光合色素含量测定
-
生产实践上,常用嫁接成活率和植株生长量等指标来评价砧木与接穗之间的亲和力. 本试验中巫山脆李嫁接在野生李上嫁接成活率最高,植株生长量最大,表现亲和,这与在绥李[14]上的研究一致. 而山樱桃和山杏砧木嫁接苗嫁接成活率低,生产上不宜作为巫山脆李的砧木应用.
-
叶片面积直接与接受光的多少成正比,因此与植株光合作用密切相关. 试验中毛桃砧木嫁接苗的叶片长、宽都是最小的,毛桃砧木嫁接苗叶片光合效率也偏低. 比叶重的大小主要由植物叶片的厚度和密度决定[15],两者的改变都会影响光的利用效率以及叶肉细胞导度的大小,最终导致光合效率的改变[16]. 本研究中的毛桃砧木嫁接苗叶片长、宽均小于其余3种,比叶鲜质量和比叶干质量小于山樱桃砧木嫁接苗的叶片,与野生李和山杏砧木嫁接苗叶片一致. 这与毛桃砧木嫁接苗叶片厚、密度大有关,与在枇杷[17]上的研究相一致.
-
植物中光合色素均参与了植物的光合作用,包括叶绿素和类胡萝卜素. 其中Chl a可以吸收、传递和转化光能[18]. Chl a和Chl b可以相互转换,其比值是植物适应不同生境的表现,其在干旱胁迫和重金属胁迫下都具有参考意义[19,20]; 类胡萝卜素可以吸收、传递电子和清除自由基,在光合作用和抗逆方面都有重要作用[21-22]. 研究表明,野生李和毛桃在干旱胁迫和重金属胁迫方面更有潜力,这与在刺茉莉[23]上的研究相符合. 不亲和砧穗组合的净光合速率、气孔导度、蒸腾速率明显低于亲和的嫁接组合,试验中毛桃的净光合速率和气孔导度显著高于其余3种嫁接组合,说明毛桃砧木与巫山脆李亲和性良好,这与在柚子[24]上的研究相一致. 叶片蒸腾作用、气孔交换和光合作用与植物的叶片结构有密切关系,影响着植株的生长发育[25],本研究中毛桃砧木嫁接苗叶片薄、组织结构疏松度高,推测这种叶片结构可能会导致毛桃砧木嫁接苗叶片在干旱胁迫下易丧失水分导致叶片萎蔫,对光能利用效率低,对干旱耐受度低,这与前人在板栗[26]、灌木[27]和黄柳[28]上的研究相一致. 本试验中山杏嫁接苗叶片栅栏组织发达,细胞层数达到了3层,栅栏组织排列紧密,叶片的组织结构紧密度最大. 因此从叶片解剖结构上判断山杏砧木嫁接苗耐旱能力最强,这与景晨娟等[29]对3种李叶片结构研究中的发现相符,表明叶片组织结构紧密程度与植株耐旱性正相关. 研究结果还显示毛桃和野生李砧木嫁接苗的叶片海绵组织厚度和组织结构疏松度要大于山樱桃和山杏砧木嫁接苗,表明毛桃和野生李耐旱能力相对于山樱桃和山杏砧木嫁接苗要弱,这与在烟草[30]叶片海绵组织结构的研究中一致,表明海绵组织厚度和叶片组织结构疏松度与植株耐旱性呈负相关.
-
嫁接口细胞学观察可以直观观察到接穗与砧木之间的愈合情况,通过硬组织切片观察发现山樱桃与巫山脆李接穗的接口处有大量木质化现象出现,木质素的沉淀导致隔离层的出现,这可能是导致山樱桃砧木和巫山脆李接穗不亲和的原因之一; 山杏砧木与巫山脆李嫁接口处有黑化现象出现,这可能是酚类物质被氧化成了有毒物质,从而导致砧穗之间出现隔离层[31],由此判断,这可能是导致山樱桃和山杏砧木与巫山脆李接穗亲和性较低的主要原因; 毛桃和野生李砧木与巫山脆李嫁接口含有大量正常的薄壁细胞,表明毛桃和野生李砧木与巫山脆李嫁接口愈合良好,这与Errea等[32]对杏嫁接在红叶李和骆建霞等[33]将苹果嫁接在海棠上的研究结果相似.
-
接穗和砧木嫁接后,嫁接苗中的SOD、POD、CAT的活性都有所提高[34-37]. 本研究中毛桃和野生李砧木与巫山脆李接穗之间愈伤组织的SOD含量比山樱桃和山杏高,毛桃砧木与巫山脆李接穗之间愈伤组织的SOD比活力比其余3种砧木中含量高,SOD可以清除植物中的超氧阴离子自由基(O2-),是植物亲和性指标之一,表明野生李和毛桃砧木与巫山脆李接穗之间亲和性良好,这与陈哲[38]在荔枝上、白雪[39]在薄皮甜瓜上的研究一致. POD是参与植物木质素合成的重要酶,其活性与木质化程度有关[40],试验结果显示山樱桃中POD活性含量最高,这也与嫁接口细胞学观察到砧穗间大量木质化现象出现相一致. 嫁接体在愈合时活性氧(ROS)含量呈现先上升后下降的趋势,这在番茄[41]中得到了证明,而CAT作为ROS清除酶,其含量也会相应跟着ROS变化. 山樱桃砧木与巫山脆李接穗之间愈伤组织的POD和CAT含量显著高于毛桃和野生李,可能是因为山樱桃砧木与接穗之间ROS含量高,表现不亲和.
-
Akira等[42]在发现拟南芥中AP2/ERF转录因子WIND1 可以通过B-ARR通路介导的细胞分裂素反应来控制细胞去分化,还可诱导油菜、番茄和烟草的愈伤组织形成[43]. 本研究中,毛桃砧木与巫山脆李接穗之间愈伤组织中WIND1 的相对表达量相显著高于山樱桃和山杏两种砧木. 嫁接亲和的组合产生的愈伤组织要比不亲和的组合更多更快,这在杏叶梨/杜梨[44]、梨/木瓜[45]和柿[46]上已经得到了证实. 毛桃和野生李砧木与巫山脆李嫁接口WIND1 的高表达于嫁接亲和相一致. 山樱桃和山杏砧木嫁接口愈伤组织的苯丙氨酸解氨酶基因PAL1 相对表达量比毛桃和野生李高,可能是因为山樱桃和山杏砧木与巫山脆李接穗之间的隔离层和酚类物质氧化导致的,这与Pina等[47]和Irisarri等[48]在杏/李嫁接组合上的研究结果相似,表明山樱桃和山杏不适宜作为巫山脆李的砧木. ARF1 是转录因子,在生长素信号传导途径中起着重要的作用,其表达量高低可以反映出嫁接口生长素含量多少,可以作为嫁接亲和性的参考指标. 本研究发现野生李砧木与巫山脆李接穗之间愈伤组织的转录因子ARF1 的相对表达量相较于其余3种砧木要高,与野生李砧木的嫁接亲和性高完全吻合,这与在砂糖橘[49]上的研究相一致. 4CL 是木质素生物合成关键酶基因,在植物生长发育过程中具有重要意义[50]. 试验结果表明,野生李砧木与巫山脆李接穗之间愈伤组织的木质素途径关键基因 4CL1、4CL2、4CL3 相较于其余3种砧木要高,表明野生李砧木与巫山脆李接穗之间木质素含量要高于其余3种砧木,这与桃[51]上的研究结果相似.
综上所述,4种砧木嫁接苗的嫁接成活率、植株生长量、叶片的净光合速率和气孔导度与砧穗亲和性显著相关,可以利用嫁接口切片观察和愈伤组织中SOD、POD酶活性以及 WIND1、PAL1、ARF1、4CL 等基因相对定量来判断砧穗亲和性高低. 综合研究表明,野生李与巫山脆李的嫁接亲和性高,嫁接成活率高,苗木长势良好,适合用于巫山脆李的砧木.
 DownLoad:
DownLoad: