





-
开放科学(资源服务)标识码(OSID):

-
家蚕是我国的重要经济昆虫,已经有5 000年的驯化历史. 近年来,家蚕成为了典型的鳞翅目模式昆虫,对家蚕的基础研究可为研究其他鳞翅目昆虫提供重要的理论支撑. 家蚕具有开放式血淋巴循环系统,组织器官浸润于血淋巴中. 血淋巴在营养物质转运、废物运输等方面发挥着至关重要的作用. 家蚕血淋巴中有几百种蛋白质,这些蛋白质主要分为2大类:营养储存蛋白和免疫相关蛋白[1]. 营养储存蛋白主要包括载脂蛋白、贮藏蛋白、卵黄原蛋白和30K蛋白. 30K蛋白是序列高度同源且分子量在30K左右的一群蛋白,在家蚕血液和胚胎中含量非常高[2-6]. 家蚕30K蛋白的功能研究将有助于我们理解30K蛋白在血液和胚胎中的作用.
2007年,SUN等[7]利用家蚕的EST数据对30K蛋白基因进行了鉴定,共鉴定到10个基因. 2012年,ZHANG等[8]利用家蚕基因组精细图序列数据,确定家蚕中共有46个基因编码30K蛋白. 根据进化分析和结构域分析,可以将30K蛋白分为3个家族:ENF肽结合蛋白、富含丝氨酸和苏氨酸的30K蛋白和典型30K蛋白. 典型30K蛋白分子量约为30 kDa,共有24个成员. 对4龄和5龄家蚕血液进行蛋白质组学分析,结果表明典型30K蛋白在5龄家蚕血液中高量存在,尤其是BmLP1和BmLP2[1]. 已有研究表明,BmLP1有多种生物学功能,在胚胎的正常发育中具有重要作用[9]. BmLP1可以与糖结合,如甘露糖、葡萄糖、N-乙酰葡糖胺[10]和葡聚糖,葡聚糖是病原体表面携带的重要糖分子,BmLP1可以结合葡聚糖,暗示其可能参与了昆虫的免疫[11-12];YE等[13]证实BmLP1可以与血细胞膜结合,通过促进血细胞包囊及黑化反应参与家蚕免疫;BmLP1可以结合EcR-B1,从而抑制蜕皮激素诱导的细胞凋亡[14]. ZHANG等[9]对BmLP7进行了详细研究,发现BmLP7主要由5龄家蚕的脂肪体合成,然后分泌进入血液中,再转运进入卵巢中,最终在胚胎发育过程中被胚胎肠腔内的酶降解,为胚胎发育提供能量和氨基酸. 近期有研究表明,BmVgR(家蚕卵黄原蛋白受体)可以与BmLP1结合,将BmLP1转运进入卵巢. 2016年,WANG等[15]从5龄家蚕消化液中纯化获得碱性丝氨酸蛋白酶P-IIc,该酶在胚胎后期表达,将该蛋白酶与不同30K蛋白孵育后,发现其具有强烈水解30K蛋白的功能.
30K蛋白成员众多,且在生物学功能上呈现出丰富的多样性. 前期研究了30K蛋白BmLP7的表达模式与功能,为了系统地比较与分析不同30K蛋白的功能差异,有必要对BmLP1等其他30K蛋白进行研究. 本研究利用半定量RT-PCR,Western blotting,免疫组化等技术,对家蚕30K蛋白BmLP1在不同组织器官中的表达模式进行调查和分析,旨在揭示家蚕BmLP1蛋白的合成、转运与利用情况,阐明家蚕30K蛋白在胚胎发育中的功能.
HTML
-
家蚕大造为本研究的实验材料,由西南大学提供. 大造采用常规饲喂方法饲养,饲喂的相对温度为25 ℃,相对湿度为60%.
Trizol试剂购于Invitrogen公司;dNTPs,Oligo (dT),M-MLV反转录酶购于Promega公司;羊抗兔二抗购于Sigma公司;Tubulin抗体、FITC染色液和抗荧光淬灭封闭液购于碧云天;DAB显色试剂盒购于北京中杉金桥;PVDF膜购于Roche;苯基硫脲、考马斯亮蓝、甘氨酸等试剂购于上海生工;松油醇、二甲苯、磷酸二氢钠、磷酸氢二钠、甲醇及冰醋酸等试剂购于重庆化学试剂厂.
-
在Silkdb数据库(
http://silkworm.swu.edu.cn/silkdb/ )下载30K蛋白BmLP1,BmLP2,BmLP3和BmLP7的氨基酸序列[16],利用ClustalX 1.83软件进行多序列比对分析[17],并利用GeneDoc软件进行手动校正. -
收集不同发育时期的家蚕脂肪体材料放于-80 ℃冰箱中保存备用. 家蚕的发育阶段共8个,分别为:5龄第3 d,5龄第5 d,5龄第7 d,上蔟第2 d,化蛹第1 d,化蛹第4 d,化蛹第7 d和化蛾第1 d. 利用Trizol试剂提取脂肪体的总RNA,反转录为cDNA. 利用cDNA为模板进行PCR扩增,扩增所用引物见表 1,扩增条件为:94 ℃预变性4 min;94 ℃变性40 s;54 ℃退火40 s;72 ℃延伸40 s;循环28次;72 ℃再延伸10 min;最后置于12 ℃保存.
-
家蚕发育至5龄第1 d时,鉴定性别,取雌性家蚕作为实验材料. 在5龄期和上蔟期,先用解剖针扎家蚕腹足,收集血液,并加入少量的苯基硫脲防止血液黑化,再取家蚕的卵巢组织存放于-80 ℃冰箱中. 在蛹期和蛾期,将家蚕固定于蜡盘上,用20 μL枪头吸取血液置于离心管中保存,再取其卵巢组织保存于-80 ℃冰箱中. 血液样品用0.1 mol/L PBS溶液稀释5倍待用. 卵巢样品采用液氮研磨的方法研磨成粉末状,加入适量的0.1 mol/L PBS溶液溶解.
利用Brandford法对血液样品和卵巢样品进行定量分析,并以牛血清白蛋白(BSA)绘制标准曲线. 根据定量结果,取2 μg蛋白质样品进行聚丙烯酰胺凝胶电泳(SDS-PAGE)分析,凝胶浓度为12.5%. 将血液和卵巢样品进行聚丙烯酰胺凝胶电泳后,通过湿转法将蛋白转移至PVDF膜上,转膜条件为200 V,转膜时间为30 min. 待转膜完成后,将膜放置在培养皿中,用5%的脱脂奶粉封闭1 h后,倒掉封闭液,加入一抗溶液孵育2 h,用TBST溶液清洗4次,再加入二抗溶液孵育2 h,再用TBST溶液清洗4次. 最后将PVDF膜浸泡在显色液中,避光反应5 min,放入曝光仪中曝光.
-
当蚕卵发育至第6 d和第10 d时,取适量的卵放入10% KOH溶液中软化,然后置于Smith固定液中24 h,接着用乙醇梯度脱水,用松油醇处理,使之变得透明. 最后,将处理好的蚕卵包埋进石蜡中,用石蜡切片机进行切片,切片厚度设置为5 μm. 石蜡切片经过脱蜡、乙醇复水、去除内源性过氧化物酶、抗原修复、封闭、一抗孵育和二抗孵育等处理后,用DAPI和DAB显色液进行显色,在显微镜下观察并拍照.
-
蚁蚕粗提物对BmLP1降解实验的反应体系如下:BmLP1的对照组中只加入了BmLP1蛋白,最终体积用缓冲液补足;蚁蚕粗提物组中只加入了蚁蚕粗提物,最终体积用缓冲液补足;BmLP1与蚁蚕粗提物孵育的实验组中加入了蚁蚕粗提物与BmLP1,其质量比为3∶1. 缓冲液为100 mmol/L的Tris-HCl,pH值为8.8. 对照组与实验组都在37 ℃下孵育2 h,保证蚁蚕粗提物中的酶与BmLP1充分反应. 反应结束后,取相同体积的量进行SDS-PAGE电泳检测降解效果.
1.1. 材料与试剂
1.2. 氨基酸多序列比对分析
1.3. 30K蛋白基因的RT-PCR检测
1.4. 30K蛋白的SDS-PAGE及Western blotting分析
1.5. 免疫组织化学定位分析
1.6. 蚁蚕粗提物对BmLP1的降解
-
已有研究表明,家蚕的典型30K蛋白在进化树上可以分为4群:Ⅰ,Ⅱ,Ⅲ和Ⅳ[8]. 其中,Ⅲ群和Ⅳ群在5龄家蚕血液中含量非常高. BmLP1与BmLP2属于典型30K蛋白Ⅲ群,而BmLP3和BmLP7属于典型30K蛋白Ⅳ群. 从silkDB网站下载的BmLP1,BmLP2,BmLP3和BmLP7的氨基酸序列可以看出,它们都有信号肽,因而都属于分泌性蛋白. 将其序列进行比对,发现BmLP3和BmLP7的序列相似性超过90%,BmLP1与另外3个30K蛋白的序列同源性不到50%(图 1).

-
已有的研究揭示了典型30K蛋白基因是在脂肪体中合成的[8-9],为了检测Bmlp1基因在脂肪体中的表达趋势,提取了不同阶段的脂肪体RNA,将其反转录为cDNA,作为后续检测的模板. 半定量RT-PCR结果表明,Bmactin3在各时期的脂肪体中表达量相当,可以用于下一步目的基因的检测. Bmlp1在家蚕5龄期和蛹前期表达量较高,在蛹后期时,表达量开始降低,这与Bmlp3和Bmlp7的表达特征相似(图 2).
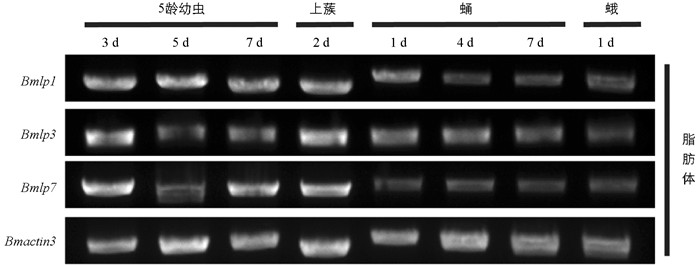
-
为了在蛋白水平上检测BmLP1的含量表达情况,利用制备的BmLP1抗体进行后续实验. 30K蛋白是一群序列结构非常相似的家族,为了检测BmLP1抗体的特异性,用已获得的原核表达蛋白进行检测. 如图 3a所示,对BmLP1,BmLP2,BmLP3和BmLP7的重组蛋白进行凝胶电泳,发现BmLP1和BmLP2的分子量比BmLP3和BmLP7略小. 利用BmLP1抗体与这4种蛋白进行孵育,发现BmLP1抗体只能识别BmLP1蛋白,表明BmLP1抗体特异性较好.
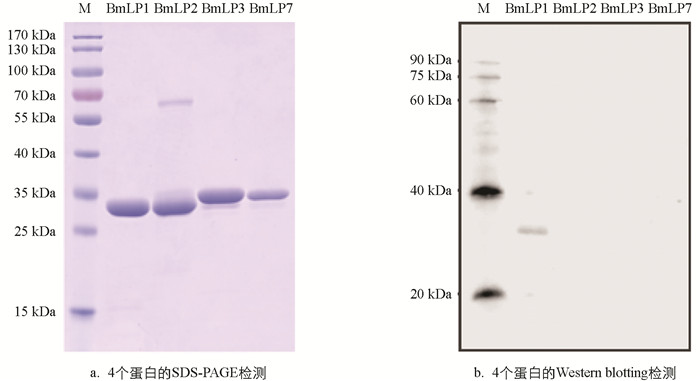
-
由于脂肪体合成的分泌性蛋白会进入到血淋巴中,为了探究BmLP1蛋白在不同时期血淋巴中的变化情况,利用BmLP1抗体进行了检测,结果如图 4所示. 由图 4可知,BmLP1蛋白在5龄期和蛹前期的家蚕血液中高量存在,而在蛾期血液中含量较少,这与脂肪体中的mRNA合成情况相似,表明BmLP1是由脂肪体合成后分泌入血液中的. 蛹后期血淋巴中的BmLP1逐渐减少,而卵巢中的BmLP1却逐渐增多,在蛾期的卵巢中含量达到最高. 而30K蛋白基因在蛹后期和蛾期的卵巢中根本不表达,这表明卵巢中的30K蛋白并非卵巢本身合成,而是由血液转运而来[9].
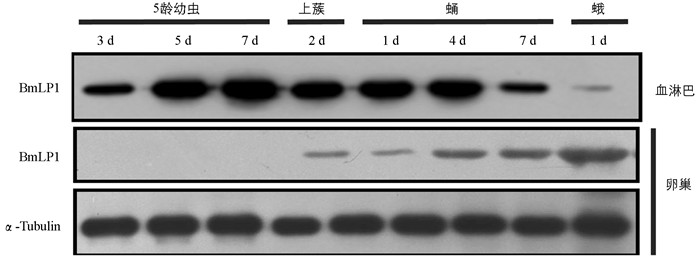
-
BmLP1蛋白从血液转运进卵巢后,一直储存在卵巢的未受精卵中,当家蚕产下卵时,BmLP1仍然存在于卵中. 为了了解BmLP1蛋白在胚胎发育过程中的变化情况,我们对第6 d和第10 d的受精卵进行了免疫组织化学定位分析,结果如图 5所示. 在卵第6 d时,胚子的雏形可见,发现BmLP1定位于胚子外边的卵黄颗粒中. 当胚胎发育至第10 d,即快孵化时,能明显地看到胚子的形态,此时,BmLP1定位于胚子的肠道内. 这表明,BmLP1最开始定位于胚外的卵黄颗粒中,但快孵化时,BmLP1被胚子吞入进了肠道内,为胚胎的孵化提供氨基酸或能量.
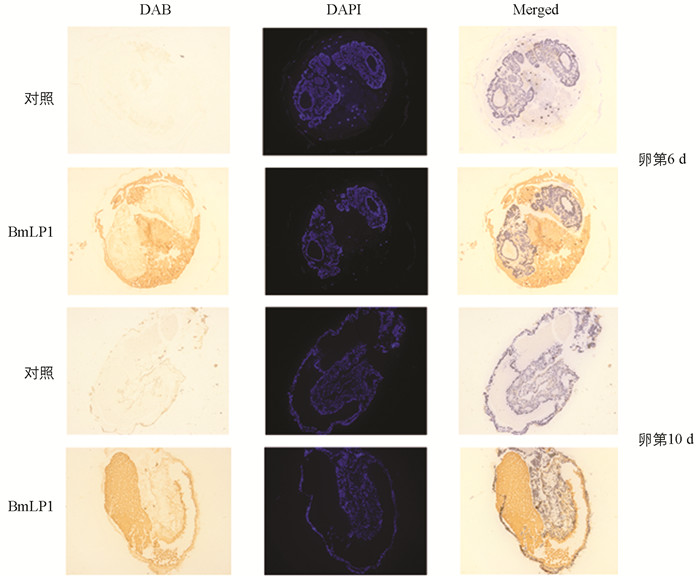
-
在胚胎发育后期,BmLP1被吞入肠腔内,猜测肠腔内有蛋白酶可以降解BmLP1,最终降解的BmLP1可以为胚胎发育提供能量或氨基酸. 为了验证这一猜测,提取蚁蚕的粗提物,将蚁蚕粗提物与BmLP1进行孵育,结果如图 6所示,蚁蚕粗提物可以将BmLP1水解.
2.1. BmLP1与其他30K蛋白的相似性分析
2.2. Bmlp1在脂肪体中的表达特征分析
2.3. BmLP1抗体的特异性检测
2.4. BmLP1在血液和卵巢中的表达情况
2.5. BmLP1在胚胎时期的定位分析
2.6. BmLP1可以被蚁蚕粗提物降解
-
在家蚕的末龄幼虫中,血液重量一般占体质量的1/4[18]. 在5龄期幼虫的血液中,30K蛋白是含量最大的一群蛋白质之一[4-5, 9]. 在其他鳞翅目昆虫,如烟草天蛾、蓖麻蚕、琥珀蚕和印度柞蚕中也鉴定到了30K蛋白的同源基因[8],但这些物种的血液中30K蛋白的表达量并不高.
本研究选取了在家蚕5龄第7 d血液中含量最高的典型30K蛋白BmLP1进行研究,发现Bmlp1基因在脂肪体中高量表达,表明BmLP1和其他典型30K蛋白均在脂肪体中合成[19]. 由于Bmlp1编码的蛋白质具有信号肽,是一种可分泌蛋白,在脂肪体中表达的Bmlp1基因分泌进入血液中,因此,在5龄期和蛹前期,血液中BmLP1蛋白含量都较高. 但在蛹后期和蛾期,BmLP1蛋白逐渐减少,与此同时,卵巢中的BmLP1却开始增加. 卵巢从蛹期开始急剧生长,卵巢管冲破卵巢膜,游离于体腔中. 卵在卵巢管中形成,并从血液中摄取营养以支持其在蛹期的快速生长. BmLP1被卵巢从血液中吸收并积累到卵中,作为重要的营养物质. 这些转运进卵巢的BmLP1在胚胎发育过程中一直不被利用,直到胚胎后期才开始减少. XU等[21]的研究表明BmLP1是通过BmVgR转运进卵巢的. 免疫组织化学定位分析结果表明,BmLP1蛋白一直储存在卵黄颗粒中,当胚子发育成熟后,胚子将含有BmLP1的卵黄颗粒吞入肠道内. 蚁蚕粗提物孵育实验结果表明,蚁蚕肠道内存在可以降解BmLP1的蛋白酶,最终将BmLP1降解为氨基酸. 这些结果与之前报道的BmLP3和BmLP7的结果相似[8],暗示了典型30K蛋白的共同特性:合成于脂肪体,分泌入血液,转运进卵巢,贮存在卵中,在蚁蚕孵化前被降解掉.
孵育实验结果表明,蚁蚕粗提物可以将大部分BmLP1水解. 2016年,WANG等[15]从家蚕的5龄幼虫肠液中纯化到一种碱性丝氨酸蛋白酶P-IIc,该蛋白酶最适pH值为11,最适温度为25 ℃. 碱性丝氨酸蛋白酶P-IIc是一个广谱性蛋白酶,可以水解大部分桑叶蛋白质. 另外,该蛋白酶在胚胎后期具有很高的表达量,可以水解卵黄磷蛋白、ESP蛋白和30K蛋白. 值得注意的是,该蛋白酶可以水解大部分BmLP1蛋白,只能部分水解BmLP2、BmLP3、BmLP7和BmLP8[15],表明其对不同底物的活性不同. 因此,认为家蚕胚胎后期,肠中的丝氨酸蛋白酶P-IIc是导致30K蛋白BmLP1降解的主要水解酶.
-
本研究发现30K蛋白Bmlp1基因在5龄幼虫和蛹期的脂肪体中高量表达,分泌进入血液中. 随着蛹期卵巢管的萌发,BmLP1逐渐从血液转运进卵巢. BmLP1蛋白一直储存在卵巢管的卵中,最终定位于卵黄颗粒中储存. 当胚子发育成熟后,胚子将含有BmLP1的卵黄颗粒吞入肠道内. 肠道内存在可以降解BmLP1的蛋白酶,最终将BmLP1降解为氨基酸,为胚胎的发育提供营养.
 DownLoad:
DownLoad: