





-
开放科学(资源服务)标识码(OSID):

-
类胡萝卜素是一类重要脂溶性色素的总称,是由异戊二烯骨架构成的C40或C30萜类化合物,也是植物中重要的次生代谢产物[1]. 类胡萝卜素主要存在于植物叶绿体以及花和果实的有色体内,是果实色泽形成的重要影响因子. 此外,类胡萝卜素作为植物激素脱落酸(ABA)等的前体物质,参与其合成代谢过程[2]. Havaux等[3]证明类胡萝卜素可以充当植物发育和对外界环境的应激反应因子,参与抵御生物和非生物逆境胁迫.
目前,类胡萝卜素代谢途径[1]已基本阐明,异戊烯焦磷酸(IPP)与二甲基丙烯焦磷酸(DMAPP)缩合,在牻牛儿牻牛儿基焦磷酸合酶(GGPPS)的作用下生成前体物质——牻牛儿牻牛儿基焦磷酸(GGPP). GGPP在八氢番茄红素合成酶(PSY)、八氢番茄红素脱氢酶(PDS)、6-胡萝卜素异构酶(ZISO)、ζ-胡萝卜素脱氢酶(ZDS)和胡萝卜素异构酶(CRTISO)等一系列酶的作用下生成番茄红素、α-胡萝卜素、β-胡萝卜素、叶黄素、玉米黄质、新黄质等类胡萝卜素,其中新黄质是ABA的合成前体物质. 苹果果实色泽丰富,有关苹果色泽的研究多集中于花青苷[4-5],而类胡萝卜素方面的研究涉及较少. 异构化作用是类胡萝卜素代谢的重要环节,CRTISO催化顺式番茄红素向全反式番茄红素转变[6]. Park等[7]在拟南芥中定位并克隆了CRTISO基因,证实了CRTISO催化多顺式类胡萝卜素异构化为全反式类胡萝卜素. 番茄tangerine(t)突变体由于CRTISO基因突变,导致原番茄红素积累,果实呈现橘粉色[8],西瓜中CRTISO基因突变也导致成熟果实中原番茄红素积累[9]. 在甜瓜中的研究表明,CRTISO可以显著提高甜瓜中的叶绿素和类胡萝卜素含量[10]. CRTISO是逆境胁迫研究中的潜在目标. 番茄中的CRTISO通过增强抗氧化系统活性和刺激次生代谢物的产生提高其对干旱的耐受性[11];在野生红球藻中CRTISO也可以显著促进虾青素和植物激素的合成[12]. 最近的一项研究结果表明小麦中TaCRTISO可以显著提高小麦的株高和单株穗数[13];水稻中的ZEBRA2编码类胡萝卜素异构酶,在水稻叶片中不仅可以促进类胡萝卜素和叶绿素的合成,还可以参与光保护,避免高光下的光氧化损伤[14],这种现象同样在芥蓝中被发现[15]. 拟南芥中CRTISO通过影响原片层体(PLBs)的生成影响叶绿体的结构,进而调节拟南芥光形态的建成和叶绿体的发育[7]. 这些研究结果证实了CRTISO在叶绿素合成以及参与光合作用中发挥重要作用,表明CRTISO在类胡萝卜素、叶绿素和逆境胁迫中具有重要的潜在功能. 在不同苹果基因型果实中类胡萝卜素含量与通路基因的表达分析表明,MdCRTISO的表达与苹果成熟果实中类胡萝卜素积累存在显著相关性[16].
苹果(Malus domestica)是世界四大水果之一,地域分布极为广泛. 中国是世界上最大的苹果生产国和消费国[17],苹果产业是主产区农业农村经济发展的支柱产业[18]. 但是,干旱、低温、盐碱等胁迫影响了苹果植株的生长发育,进而影响苹果产量和品质,对我国苹果产业产生了严重影响. 挖掘抗逆基因并解析其生物学功能对苹果产业发展具有重要意义. 前期研究发现,苹果果实中MdCRTISO的表达显著影响类胡萝卜素和ABA的合成[16],但苹果MdCRTISO基因的特性及其对逆境的响应模式尚未见报道. 因此,本研究从金冠苹果叶片中克隆MdCRTISO基因,采用生物信息学方法对其编码蛋白的理化性质、保守功能域、亲/疏水性、空间结构及系统发育关系等进行分析,并利用实时荧光定量(RT-qPCR)分析MdCRTISO基因在苹果不同组织和不同胁迫条件下的表达模式,旨在为后续深入研究该基因的功能以及通过基因工程手段培育优良苹果品种奠定理论基础.
HTML
-
本氏烟草(Nicotiana benthamiana)用于亚细胞定位,组织特异性分析采用富平楸子(Malus prunifolia)的不同组织,包括根、叶、茎、花和果实. PEG处理采用长势一致、真叶6~7片的平邑甜茶(M. hupehensis)水培苗,用含20% PEG 6000的1/2 Hoagland营养液进行处理,之后分别在处理0 h、6 h、12 h、24 h采样. 盐胁迫处理挑选长势一致、真叶6~7片的平邑甜茶水培苗,用含200 mmol/L NaCl的1/2 Hoagland营养液进行处理,之后分别在处理0 h、6 h、12 h、24 h采样. ABA处理挑选长势一致、真叶6~7片的平邑甜茶水培苗,用含100 μmol/L ABA的1/2 Hoagland营养液进行处理,之后分别在处理0 h、6 h、12 h和24 h采样. 低温处理采用生根移栽30 d后的嘎啦-3(M. domestica,GL-3)组培苗,在4 ℃条件下低温处理,分别在处理0 h、6 h、12 h和24 h取样. 碱胁迫处理挑选长势一致、真叶6~7片的平邑甜茶幼苗用1/2 Hoagland营养液(用H2SO4调pH值为6.0±0.1)进行水培,预培养2周后,用1 mol/L的Na2CO3∶NaHCO3=1∶1(v∶v)将营养液pH值调到9.0±0.1进行碱胁迫处理,之后分别在处理0 h、6 h、12 h和24 h采样. 所有材料取材后液氮速冻,置于-80 ℃保存备用.
-
以金冠苹果叶片为材料,采用改良的CTAB法提取样品RNA. 然后用一步RT-qPCR试剂盒(购自上海生工生物技术有限公司)反转录合成cDNA. 利用Primer Premier 5.0设计引物,MdActin作为内参基因[16](表 1). 使用Light Cyler实时PCR仪器(Roche Diagnostics Corporation,Indianapolis,IN,USA),RT-qPCR试剂为SYBRⒸ Premix Ex Taq TM Ⅱ J. 10 μL体系如下:引物F(10 μmol/L) 0.4 μL,引物R(10 μmol/L) 0.4 μL,SYBR Premix 5 μL,cDNA 1 μL,ddH2O 3.2 μL. PCR反应条件:95 ℃预变性30 s;95 ℃变性10 s,58 ℃退火30 s,72 ℃延伸10 s,40个循环;37 ℃ 30 s. 通过2-ΔΔCT方法计算基因的相对表达水平. 每个处理3次生物学重复.
-
将提取的金冠苹果叶片RNA反转录成cDNA作为模板,然后用CRTISO的特异性引物进行扩增(表 1). PCR反应体系为25 μL,模板1 μL,上下游引物(10 μmol/L)各1 μL,PrimeSTAR Mix高保真酶12.5 μL,无菌水9.5 μL. PCR反应条件为95 ℃变性5 min;98 ℃ 10 s,53 ℃ 15 s,72 ℃ 30 s,进行35个循环;72 ℃延伸5 min. 扩增产物用琼脂糖凝胶电泳检测.
-
在NCBI下载CRTISO蛋白序列,使用Expasy server(
http://www.expasy.org/tools/ )的ProtParam和ProScale工具分析蛋白质的分子量、等电点、亲/疏水性、脂肪族指数等. 利用SWISS models(https://swissmodel.expasy.org/ )进行蛋白质三级结构预测,利用NCBI的CDD数据库进MdCRTISO蛋白保守结构域预测. 利用PlantCare网站(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )工具分析启动子序列顺式作用元件,并对顺式作用元件可视化. -
利用植物基因组网站Phytozome(Ⅴ13)(
https://phytozome-next.jgi.doe.gov/ )BLASTP工具,基于目标基因蛋白序列在梨(Pyrus spp.)、梅(Prunus mume)、桃(Prunus persica)、欧洲甜樱桃(Prunus avium)、拟南芥(Arabidopsis thaliana)和玉米(Zea mays)等不同物种基因组中进行同源序列检索并下载相应序列,使用DNAMAN软件引进行序列相似性比对,并利用MEGA 7.0软件构建系统进化树. -
将MdCRTISO基因扩增产物连接到pCAMBIA2300-GFP载体上,转化大肠杆菌DH5α,测序正确后提取质粒,并转化农杆菌GV3101. 将检测正确的菌液接种于YEP液体培养基培养12 h,再大摇12 h至OD600为1.0,离心后用10 mmol/L MES重悬液重悬菌体,使OD600为0.8~1.0. 注射到烟草叶片中,侵染48 h后,使用激光共聚焦显微镜(TCSSP8-SR,Germany)观察GFP荧光.
1.1. 试验材料
1.2. 试验方法
1.2.1. RNA提取及RT-qPCR
1.2.2. MdCRTISO基因CDS的克隆
1.2.3. 生物信息学分析
1.2.4. 同源性分析
1.2.5. 亚细胞定位
-
以反转录金冠苹果叶片cDNA为模板,PCR扩增CRTISO基因的全长序列,如图 1a所示,琼脂糖凝胶电泳显示在2 100 bp左右有清晰、特异的条带. MdCRTISO基因序列号为MD14G1044400(LOC103454192),序列分析结果表明,cDNA序列全长为2 028 bp,预测蛋白分子量为74.331 kDa,原子总数10 473个,等电点pI为8.64,不稳定系数36.82,脂肪族指数为89.21,带负电荷的残基总数67,带正电荷的残基总数75,编码675个氨基酸. 蛋白质三级结构预测发现MdCRTISO蛋白包含螺旋、折叠、卷曲等二级结构(图 1b),且MdCRTISO是稳定的亲水性蛋白(图 1c). 根据氨基酸序列分析的结果,MdCRTISO具有一个保守的结构域位于N端的第150至675个氨基酸之间(图 1d),分别是八氢番茄红素脱氢酶相关蛋白(次级代谢物的生物合成、运输和分解代谢)和胡萝卜素异构酶(辅因子、假体组和载体的生物合成及其他).
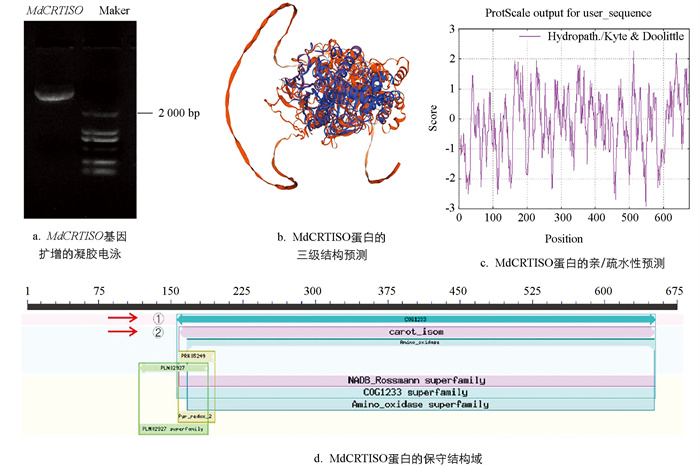
对MdCRTISO基因启动子进行顺式作用元件分析,发现该启动子序列上包含多种不同功能顺式作用元件(表 2). 与植物激素相关的顺式作用元件包括:ABRE、TGA-element、CGTCA-motif、TGACG-motif;与逆境相关的顺式作用元件包括:ARE、AT-rich element;同时,还有大量光响应相关元件如AE-Box、G-Box、GATA-motif和MRE.
-
将MdCRTISO蛋白与其他物种CRTISO蛋白进行氨基酸序列多重比对,结果如图 2所示,苹果MdCRTISO蛋白与其他物种CRTISO蛋白序列整体相似度为76.86%,说明这些蛋白在进化上非常保守,暗示它们在植物进化过程中发挥重要的作用. 其中,与梨、梅、欧洲甜樱桃、桃的相似度分别为90.32%、83.54%、83.13%、81.88%,表明它们可能具有相似的功能.
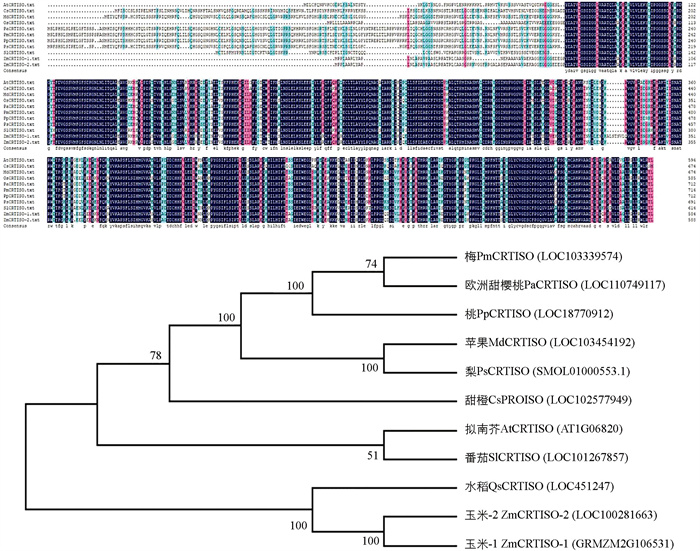
基于10个植物来源CRTISO蛋白构建的系统进化树,如图 2所示. 11个参试物种CRTISO基因编码蛋白经聚类可分为两大分支,玉米和水稻被单独分为一类,苹果和其他物种被聚在另外一个分支. 其中苹果MdCRTISO与梨的PsCRTISO亲缘关系最近,桃、欧洲甜樱桃和梅次之;甜橙的CsCRTISO也与MdCRTISO保持较高的亲缘关系;拟南芥和番茄的CRTISO在同一个分支且与苹果MdCRTISO保持较高亲缘关系,而苹果MdCRTISO与单子叶植物水稻和玉米CRTISO的同源性较远.
-
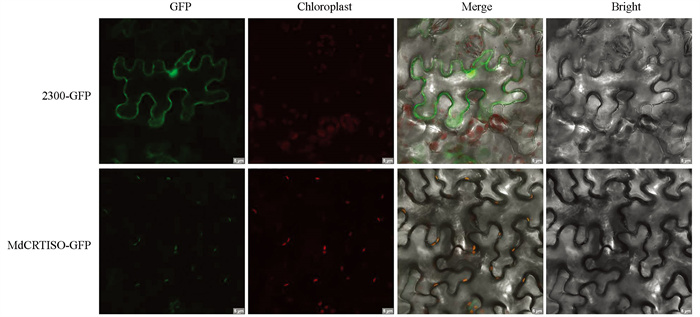
在本氏烟草中瞬时表达MdCRTISO蛋白,分析该蛋白的亚细胞定位情况. 由图 3可以看出,GFP对照在整个烟草叶片细胞,包括细胞质、细胞核、细胞质膜上都能观察到荧光,而瞬时过表达MdCRTISO烟草叶片仅在叶绿体上观察到荧光,表明MdCRTISO蛋白定位于叶绿体.
-
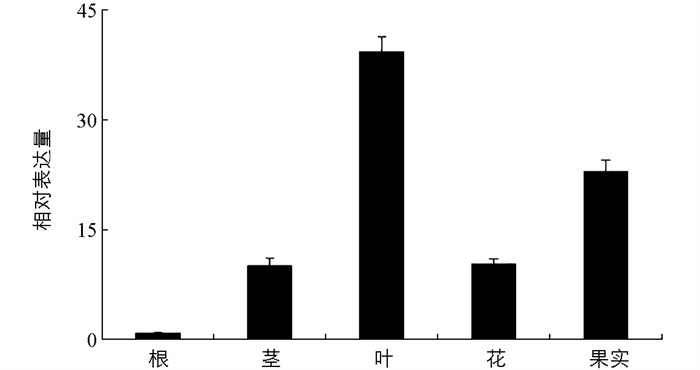
采用RT-qPCR分析MdCRTISO基因在苹果不同组织中的相对表达量. 结果表明,MdCRTISO在苹果根、茎、叶、花和果实中均有表达,其中在果实和叶片中表达较高,在花和茎中表达次之,在根中表达最低(图 4).
-
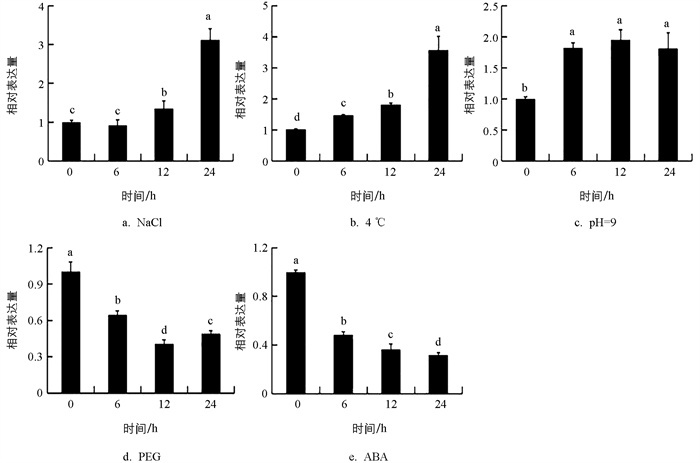
MdCRTISO启动子上含有多个与逆境相关的顺式元件,类胡萝卜素作为ABA的合成前体,相关研究表明过表达该基因显著提高ABA含量[16],为探究MdCRTISO基因对不同胁迫的响应,对苹果在盐碱、低温、干旱等多种胁迫下的表达响应模式进行了分析. 研究结果表明,MdCRTISO在12 h和24 h均能显著响应盐胁迫,盐胁迫24 h时其表达量上调到3.12倍. MdCRTISO在6 h、12 h和24 h均能显著响应低温胁迫,低温胁迫24 h其表达量上调到3.56倍. MdCRTISO在6 h、12 h和24 h均能响应碱胁迫,碱胁迫12 h时其表达量达到最高值,为0 h的1.95倍. MdCRTISO在6 h、12 h和24 h均能显著响应干旱胁迫,干旱胁迫12 h时其表达量达到最低值,仅为0 h的41%. MdCRTISO在6 h、12 h和24 h均能显著响应ABA胁迫,ABA胁迫24 h时其表达量下调到0 h的30%(图 5). 以上结果表明MdCRTISO可能参与盐、低温、碱、干旱和ABA胁迫的响应.
2.1. MdCRTISO基因克隆与生物信息学分析
2.2. MdCRTISO蛋白的同源比对和系统进化树分析
2.3. MdCRTISO亚细胞定位分析
2.4. MdCRTISO在苹果不同组织中的特异性表达分析
2.5. MdCRTISO在不同逆境下的响应分析
-
类胡萝卜素不仅是决定果实营养价值的重要指标,作为果实的主要呈色色素,同时也是影响果实外观品质的重要因素. 研究表明类胡萝卜素可以通过清除植物体内的活性氧来降低氧化胁迫对植物造成的伤害[19]. 随着类胡萝卜素合成代谢研究的深入,植物类胡萝卜素代谢途径中的酶和基因已被广泛研究. 例如,从柚转录组数据筛选得到CmPSY、CmPDS、CmZDS和CmLCYB等参与蜜柚类胡萝卜素合成途径关键酶基因的序列,并进行了表达分析[20]. 香蕉中负责编码类胡萝卜素羟化酶CYP97A3的LUT5 (Lutein deficient 5)基因,在果肉类胡萝卜素途径中起正调控作用,能促进类胡萝卜素形成[21]. 苹果果实中,PSY1过表达增加了类胡萝卜素含量,且PSY1过表达在一定程度上可减少套袋避光对类胡萝卜素含量的影响[22]. 这些研究表明类胡萝卜素在果实品质与逆境响应中具有很好的应用潜力.
CRTISO是催化顺式番茄红素向全反式番茄红素转化的关键酶. 番茄中SlCRTISO在叶片和成熟果实中表达水平最高,且在叶片中胡萝卜素的含量与CRTISO-L1和CRTISO-L2的表达水平呈正相关[23],这些结果同样在烟草和苹果中得到了验证[16, 24-25],表明CRTISO是类胡萝卜素生物合成的关键酶,且CRTISO可以显著促进苹果类胡萝卜素和ABA的合成. 柑橘CRTISO基因的表达能够恢复柑橘突变体类胡萝卜素的生物合成[26]. 三角褐指藻中的研究表明CRTISO基因表达量与岩藻黄素含量呈线性关系,表明三角褐指藻岩藻黄素的生物合成是通过调控CRTISO基因的表达来实现的[27]. 同样,枸杞中的LcCRTISO过表达显著增加了转基因烟草中类胡萝卜素的积累,增强了烟草光合作用和对盐胁迫的耐受性[25],表明LcCRTISO可能通过影响类胡萝卜素的合成间接影响植物的光合作用,进而提高植物对胁迫的耐受性;在芥蓝中敲除CRTISO基因会导致其叶绿素和类胡萝卜素的含量大幅下降,导致其对逆境胁迫的耐受性降低[15];水稻中的CRTISO已经被证明显著影响光合作用[14]. 这些研究结果显示CRTISO作为潜在因子在类胡萝卜素合成、ABA合成、叶绿素合成和光合作用中起到重要功能. 本研究在GL-3苹果幼苗中外源施加ABA,分别在6 h、12 h、24 h发现CRTISO表达受到显著抑制,这种抑制可能是ABA的过量引起的反馈调节导致,这个结果与CRTISO可以显著促进ABA合成的结果是一致的[16];此外,本研究还发现MdCRTISO的表达显著受到NaCl和4 ℃处理的诱导,表明MdCRTISO响应低温和盐胁迫,推测在盐胁迫和低温胁迫存在的情况下,机体的免疫调控机制诱导ABA的增加和光合作用的增强来抵御胁迫环境,而这种机制的实施需要CRTISO的超量表达来实现,具体的分子机制需要更加深入的研究. 这些研究结果均表明CRTISO是提高苹果对盐和低温处理的潜在因子. 有趣的是,在PEG处理6 h、12 h、24 h条件下,MdCRTISO表达均下调,这可能是受到不同的调节机制所致. 以上研究均表明CRTISO是类胡萝卜素的生物合成中的关键限速酶,且该基因在提高植物对胁迫的耐受性中发挥潜在的功能,这为植物类胡萝卜素与逆境胁迫协同研究提供了一定参考.
RT-qPCR分析表明MdCRTISO基因在苹果根、叶、花和果实等各组织中均有表达,但在叶和果实中高度表达,暗示MdCRTISO在苹果不同组织部位可能具有不同的生物学功能. MdCRTISO在苹果叶片中的高表达表明CRTISO在叶片光合作用以及类胡萝卜素——叶绿素稳态中可能发挥更重要的作用. MdCRTISO在苹果果实中的高表达表明CRTISO在果实类胡萝卜素合成中具有重要功能.
启动子顺式作用元件是基因功能的重要组分,反映基因潜在的功能和调控途径. 本研究中,苹果MdCRTISO基因启动子的顺式作用元件可分为激素响应、胁迫响应和生长发育相关等3类,其中有大量响应生长发育和逆境相关的元件(如ABRE、TGA-element、ARE、AT-rich element),暗示该基因在植物生长发育和逆境胁迫响应中可能发挥着不同的作用. 本研究发现在盐胁迫和冷胁迫下MdCRTISO转录水平均显著上调,在ABA处理和干旱胁迫下均显著下调,这些结果初步说明,MdCRTISO基因可能受多种逆境胁迫诱导,具体分子机制有待进一步研究.
本研究利用生物信息学方法分析MdCRTISO基因结构、理化性质、顺式元件和系统进化等,并检测了MdCRTISO基因在不同组织和不同逆境胁迫下的表达,为苹果MdCRTISO基因的功能研究奠定了一定的科学基础,关于MdCRTISO在苹果类胡萝卜素积累和逆境响应中的功能及其分子调控机制将有待进一步研究.
 DownLoad:
DownLoad: