





-
开放科学(资源服务)标识码(OSID):

-
埃塞俄比亚芥菜属于十字花科芸薹属作物,由黑芥和甘蓝天然杂交,自然加倍进化而来[1-3]。埃塞俄比亚芥菜的幼苗生长旺盛,并且具有抗寒、抗虫、抗旱、抗裂角等优异性状,具有较大的生产应用前景[4-6]。埃塞俄比亚芥菜的种子可用作调味品(芥末),也可用于制作药品、化妆品、塑料等。在育种中,可将埃塞俄比亚芥菜与现有品系进行杂交,转移单个优异性状,目前已成功转移黄籽基因、抗黑胫病基因、抗白粉病基因等[7]。通过埃塞俄比亚芥菜和芸薹属其他作物杂交会产生一些中间多倍体品系,利用这些多倍体品系可对目标性状进行改良,从而丰富遗传多样性[8-11]。通过将白菜型油菜的A基因组(Ar)与埃塞俄比亚芥菜的C基因组(Cc)进行种间杂交,成功培育出具有显著生物量和产量优势的新型甘蓝型油菜(ArCc)。研究显示,随着外源ArCc含量的增加,这些优势呈现出增强的趋势[12-13]。构建芸薹属异源六倍体(ArArBcBcCcCc) 的种质库,运用高通量技术比较了人工合成芸薹属六倍体与其亲本之间的转录组差异,共得到35 644 409条清洁读段,匹配到的基因数为32 642,其中在父本白菜型油菜、母本埃塞俄比亚芥菜和六倍体植物中分别有29 260、29 060和29 697个基因被检测到[14]。通过对异源六倍体的染色体稳定性、基因组稳定性和育性的分析表明,异源六倍体比例和稳定性随着自交代数的增加而增强,且花粉育性接近正常[15]。利用该异源六倍体种质库与甘蓝型油菜杂交产生五倍体(ArAnBcCcCn),自交多代产生新型甘蓝型油菜,与传统甘蓝型油菜相比,新型甘蓝型油菜具有更丰富的变异[15]。然而埃塞俄比亚芥菜这些优良特性的利用受限于其硫苷含量较高,通过对埃塞俄比亚芥菜品质进行改良,可满足人们对其食用品质、饲用品质以及工业用途的不同需求。
硫苷是一组含氮、硫的植物次生代谢物,对植株的长势、耐旱性、广谱抗病性等方面具有显著影响。在十字花科芸薹属作物中共发现了约30种硫苷[16]。硫苷及其降解产物具有抵御草食动物、抗病虫及抗病原微生物的能力,在植物的防卫反应中发挥重要的作用。如硫苷降解产物烯丙基异硫氰酸酯、2-苯乙基异硫氰酸酯等对芸苔属专食性昆虫具有致死作用。当植株中2-苯乙基硫苷含量增加时,蚜虫数量减少,植株抗虫性增强。此外,硫苷还参与调节植物的耐旱性,在干旱胁迫下,某些油菜品种的硫苷含量会发生变化,而这种变化可能与植物的抗旱机制相关联。然而,对于某些作物如油菜而言,过高的硫苷含量会影响其品质,如硫苷及其降解产物是油菜籽饼粕中主要的抗营养因子,会影响菜籽饼粕的饲用安全。因此,降低硫苷含量成为油菜品质育种的一个重要目标。油菜种子中的硫代葡萄糖苷,由于其水解产物的毒性被认为具有抗营养作用。2-羟基-3-丁烯基硫代葡萄糖苷酸酯转变为促甲状腺激素产物,而3-丁烯基和4-戊烯基硫代葡萄糖苷可以转变为异硫氰酸酯,这会降低牛、猪和家禽对饼粕的适口性[17]。研究者通过RNAi抑制硫代葡萄糖苷路径中R2R3-MYB转录因子MYB28的表达来生产低硫苷的芥菜型油菜品种,该基因是脂肪族硫代葡萄糖苷生物合成的主要调节剂[18]。此外,通过基于乙基磺酸盐的TILLING方法破坏硫苷转运蛋白可获得低硫苷白菜型油菜[19]。但是,该方法难以应用于具有多基因拷贝的埃塞俄比亚芥菜品种。研究发现,埃塞俄比亚芥菜种子富含硫代葡萄糖甙和芥酸、油酸等多种脂肪酸,相对于低硫苷的甘蓝型油菜,埃塞俄比亚芥菜种子中硫代葡萄糖苷的含量仍然很高,其中丙烯基硫苷占种子总硫苷含量的95%以上[20]。大量的链式烯基硫代葡萄糖苷严重影响了埃塞俄比亚芥菜饼粕作为动物饲料的使用,研究者通过尝试化学诱变和种间杂交等方法减少硫苷特别是种子中链式烯基硫代葡萄糖苷的含量,但由于硫代葡萄糖苷复杂的生物合成过程和相关的遗传控制,收效甚微。
本研究通过克隆及数据库比对共得到6个BcMYB28同源基因,对BcMYB28同源基因的序列特征进行分析,利用CRISPR/Cas9创制BcMYB28基因敲除植株,采用代谢组分析BcMYB28敲除植株的代谢组分变化,结合转录组测序和qRT-PCR筛选埃塞俄比亚芥菜中硫苷合成途径的关键基因。研究结果为低硫苷埃塞俄比亚芥菜品质改良提供了理论依据。
HTML
-
试验材料为埃塞俄比亚芥菜种子,由西南大学油菜工程中心提供。选取埃塞俄比亚芥菜籽粒饱满的种子,用纸盘发苗,温度在22~25 ℃,浇足水分,暗光培养2~3 d,以减少幼苗的徒长现象,待种子萌发后再移到光照培养室;幼苗长出4片叶子时,选取单株嫩叶,采用CTAB法提取DNA,紫外分光光度计检测每个单株的DNA浓度,并稀释至50 ng/μL,于-20 ℃保存备用。采用TA克隆的方法获得目的基因序列信息[21]。
采用Geneious软件进行基因序列比对,利用MEGA6.0软件分析目的基因与十字花科芸薹属其他作物同源基因的亲缘关系,并构建系统进化树。
-
取埃塞俄比亚芥菜籽粒饱满的种子,用纸盘发苗,待长出4片叶子后移入土壤中,放于光照培养箱培养。光照培养箱的生长条件为:光培16 h/暗培8 h,温度为22 ℃,光照强度为10 000 Lx。取埃塞俄比亚芥菜授粉后10 d、20 d、30 d的种子(编号WT-10DAF、WT-20DAF、WT-30DAF),设置3个生物学重复,取样后放入-80 ℃冰箱备用,采用NEB柱式小量植物总RNA抽提试剂盒提取。利用Nanodrop 2000对RNA的浓度和纯度进行检测,琼脂糖凝胶电泳测RNA完整性,Agilent 2100测定RIN值。单次建库要求RNA总量为大于10 μg,浓度≥100 ng/μL,OD260/280=1.8~2.2,OD260/230≥2.0,RIN≥6.5,28S∶18S≥1.0。将RNA进行冻干处理后,送往上海美吉生物医药科技有限公司进行高通量测序。以甘蓝型和芥菜型油菜作为参考种,对埃塞俄比亚芥菜的RNA进行测序比对分析。
-
采集WT-10DAF、WT-20DAF、WT-30DAF新鲜组织,用液氮冷冻后,保存于-80 ℃超低温冰箱备用。待所有不同发育时期的样品取样完成后,将各组织材料反转录合成的cDNA第一链进行qRT-PCR分析,qRT-PCR反应体系为20 μL,包含10 μL 2× Promega GoTaq© qPCR Master Mix、1 μL 20 μmol L1正向引物、1 μL 20 μmol L1反向引物、100 ng cDNA第一链,添加ddH2O至20 μL。使用StepOnePlusTM Real-Time PCR Systems (Applied Biosystems)进行扩增,反应程序为94 ℃ 2 min;94 ℃ 3 s,60 ℃ 30 s,40循环;60~95 ℃,熔解曲线分析。使用2-ΔΔCt法计算基因的表达量[22]。
-
对埃塞俄比亚芥菜硫苷生物合成路径中关键调节因子的候选基因设计靶点(
http://skl.scau.edu.cn/targetdesign/ ),构建CRISPR/Cas9双元载体,方法按照Ma等[23]CRISPR/Cas9载体构建方法将得到的重组载体用液氮冻融法转化农杆菌菌株GV3101,构建载体所用引物包括通用引物信息(表 1)。以子叶为外植体,转化埃塞俄比亚芥菜。 -
选择埃塞俄比亚芥菜和T1代遗传转化株授粉后的种子(分别编号CR、WT),用液氮速冻后,将样品送至上海美吉生物医药科技有限公司完成UHPLC-MS测定。采用AB Sciex Triple TOF 5600高分辨质谱仪与Agilent 1290超高效液相色谱仪。ESI离子源参数设置如下:雾化气压(GS1)60 Pa,辅助气压60 Pa,气帘气压35 Pa,温度650 ℃,喷雾电压+5 000 V(正离子模式)。流动相包括溶剂A(25 mmol/L醋酸铵和25 mmol/L氨水)和溶剂B(乙腈)。将上机的代谢物参数矫正和比对之后再与数据库进行比对和物质鉴定,用ESI+模式进行。
1.1. 埃塞俄比亚芥菜DNA提取及序列分析
1.2. RNA提取与文库构建
1.3. qRT-PCR荧光定量分析
1.4. CRISPR/Cas9载体构建及埃塞俄比亚芥菜的遗传转化
1.5. 非靶向代谢物测定
-
本研究从埃塞俄比亚芥菜的DNA材料中克隆BcMYB28基因,所用引物M28-F/-R见表 1,利用PCR扩增及TA克隆获得埃塞俄比亚芥菜BcMYB28的基因序列,将该序列提交至埃塞俄比亚芥菜测序数据库(该数据库由Agricultural & Agri-Food Canada,AAFC提供),通过序列比对分析,埃塞俄比亚芥菜中共得到6个BcMYB28同源基因,分别为BcMYB28.1、BcMYB28.2、BcMYB28.3、BcMYB28.4、BcMYB28.5、BcMYB28.6。在BRAD(
http://brassicadb.org/brad/ )及NCBI(https://www.ncbi.nlm.nih.gov/ )数据库下载获取芸薹属作物MYB28同源基因的家族成员共17个,进一步将BcMYB28.1~BcMYB28.6与同源基因进行相似度分析(表 2),结果表明,BcMYB28.1、BcMYB28.3、BcMYB28.4、BcMYB28.5、BcMYB28.6分别与BoMYB28.3、BoMYB28.2、BoMYB28.1、BniMYB28.2、BniMYB28.1的相似度最高,相似度分别为95.8%、92.2%、95.5%、93.2%、95.0%;而BcMYB28.2与各同源基因的相似度在74.6%~84.9%。该结果证明BcMYB28.1、BcMYB28.3、BcMYB28.4可能起源于C染色体组的甘蓝,而BcMYB28.5、BcMYB28.6可能起源于B染色体组的黑芥。将拟南芥AtMYB28、17个已知的芸薹属作物中MYB28同源基因以及埃塞俄比亚芥菜中6个BcMYB28共24个同源基因进行进化分析(图 1)。BcMYB28.1与BcMYB28.2聚为第一大类,且BcMYB28.1与甘蓝型油菜BnMYB28.1和甘蓝中BoMYB28.3基因聚为同一亚类;BcMYB28.4与BcMYB28.6聚为第2大类,且BcMYB28.4与甘蓝中BoMYB28.1基因聚为同一亚类,BcMYB28.6与芥菜型油菜BjMYB28.1和黑芥中BniMYB28.1基因聚为同一亚类;BcMYB28.3与BcMYB28.5聚为第3大类,该结果与序列分析结果相一致。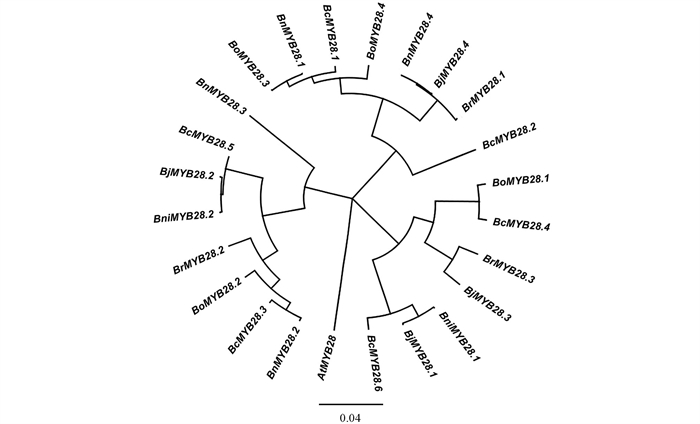
-
根据埃塞俄比亚芥菜中BcMYB28各同源基因的序列信息,通过网站(
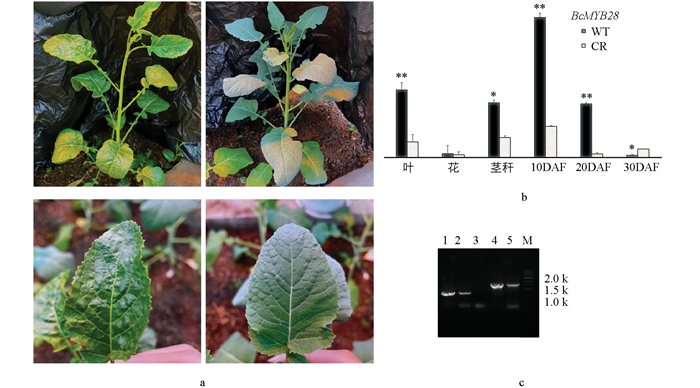
http://skl.scau.edu.cn/targetdesign/ )设计BcMYB28基因靶点T1-F/-R和T2-F/-R,构建CRISPR/Cas9双元表达载体。为确保BcMYB28.1-BcMYB28.6同时被敲除,在BcMYB28保守区域第一外显子处距ATG起始位点23 bp处和89 bp处分别设计两个靶点,通过Overlapping PCR将两个表达盒与AtU3d启动子串联,最后连入pYLCRISPR/Cas9PubiN表达载体,将构建好的重组载体pYLCRISPR/Cas9PubiN-MYB28送测序,以确保构建完整无误的双靶点载体pYLCRISPR/Cas9PubiN-MYB28。构建好的重组载体pYLCRISPR/Cas9PubiN-MYB28通过子叶为外植体的遗传转化方法转化埃塞俄比亚芥菜材料(图 2)。用引物M28-F/-R检测共得到2株T0代阳性遗传转化株(图 3c),收获T1代种子。将T1代转化植株CR与埃塞俄比亚芥菜WT种到西南大学10号温室。与WT相比(图 3a右),CR叶片呈现表面光亮、无蜡质层、无刺毛等表型(图 3a左)。取WT、CR生长期的茎秆、叶、花以及开花后10 d、20 d、30 d的种子,通过qRT-PCR分析结果显示,WT中BcMYB28在叶片、茎秆及种子形成早中期均有高量表达,CR植株中BcMYB28的表达显著低于WT(图 3b)。
-
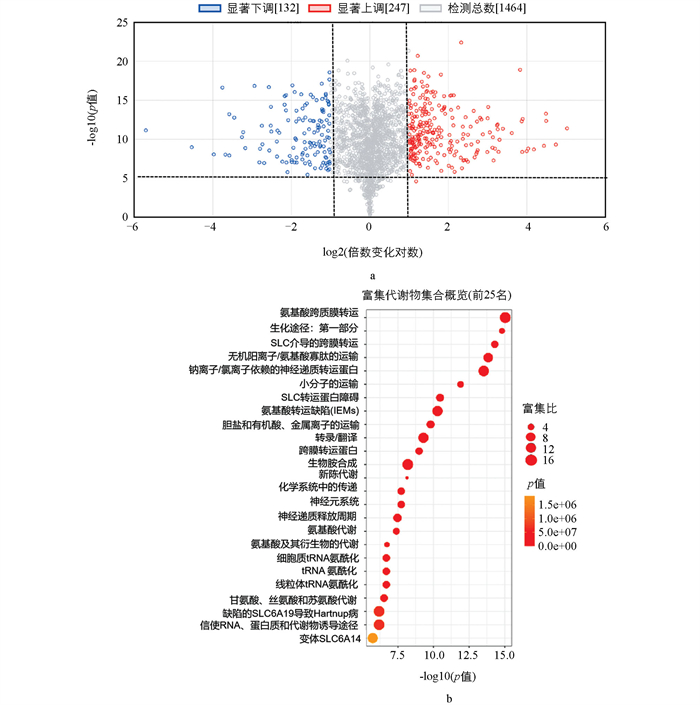
为进一步研究埃塞俄比亚芥菜中BcMYB28转录因子在硫苷生物合成路径中的作用,寻找在埃塞俄比亚芥菜种起重要作用的关键代谢物,从T1代遗传转化株、埃塞俄比亚芥菜授粉后10 d、20 d、30 d的种子材料中共鉴定出1 843种次级代谢物(表 3)。在正向模式下,与埃塞俄比亚芥菜WT相比,CR有247个代谢物含量上调,132个代谢物下调,其余1 464个代谢物无显著差异(图 4a);进一步对差异代谢物进行富集分析,找到前25个显著富集的差异代谢,甘氨酸、丝氨酸和苏氨酸代谢(Glycine,serine and threonine metabolism)被显著富集(图 4b),硫代葡萄糖苷的生物合成中半胱氨酸、丝氨酸具有重要作用,甘氨酸与苏氨酸也可对代谢产生影响。
-
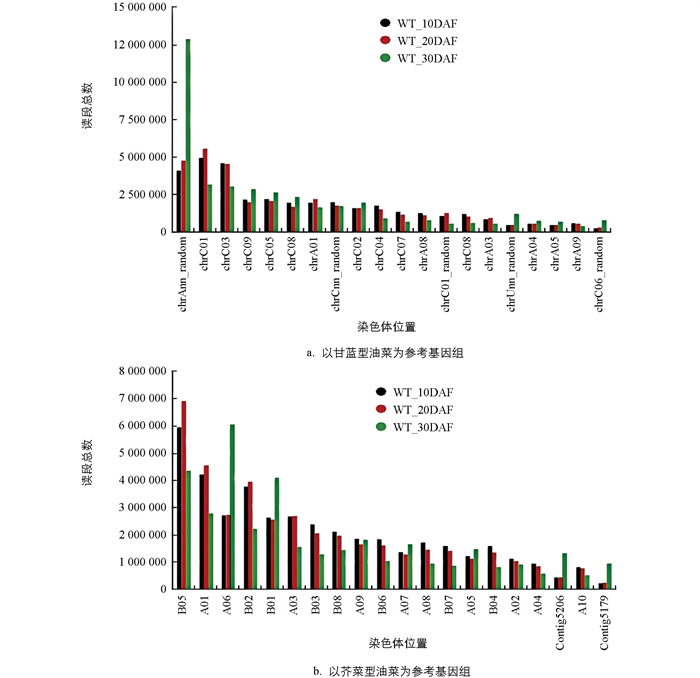
为鉴定埃塞俄比亚芥菜中BcMYB28在硫苷生物合成路径中的调节作用并找到更多埃塞俄比亚芥菜硫苷代谢中的关键调节因子,选取埃塞俄比亚芥菜开花后10 d、20 d、30 d形成期种子编号WT-10DAF、WT-20DAF、WT-30DAF(图 5),然后进行转录组测序分析。WT-10DAF、WT-20DAF、WT-30DAF中分别得到50644704、49413212、42434056条清洁读段,所有样品的错误率低于0.03%,质量为Q20(正确碱基识别率大于99%)和Q30(正确碱基识别率大于99.9%)的清洁读段分别大于98%和94%。WT-10DAF、WT-20DAF、WT-30DAF分别只有55.21%、54.24%和51.06%的清洁读段比对至甘蓝型油菜参考基因组;分别有69.6%、70.21%和72.2%的清洁读段比对至芥菜型油菜参考基因组(表 4)。埃塞俄比亚芥菜(BBCC,n=34)的清洁读段分别比对到B01-B08以及C01至C09的参考染色体上。C01和C03在C染色体组中的比对最高(图 6a),而B05在B染色体中的比对率最高(图 6b)。这可能归因于埃塞俄比亚芥菜的染色体组构成主要源自B、C组,而甘蓝型油菜则来自A、C组,芥菜型油菜则仅包含A、B组染色体。因此,埃塞俄比亚芥菜样品的清洁读段在比对时,其比对率为51.06%~72.2%。此外,当以芥菜作为参考基因组时,其比对率普遍高于以甘蓝型油菜为参考基因组的比对结果。

-
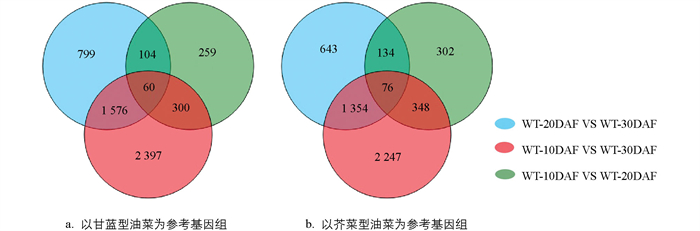
为了解埃塞俄比亚芥菜形成期种子中差异基因的表达情况,使用DESeq2对WT-10DAF、WT-20DAF、WT-30DAF分别检测差异表达基因。以甘蓝型油菜为参考物种,共鉴定出5 495个DEG,3个时期均差异表达的基因有60个(图 7a)。在每个阶段(WT-10DAF、WT-20DAF、WT-30DAF)下调表达的基因远超过了上调表达的基因,分别是1 686∶853、464∶259、3 447∶886;使用芥菜型油菜作为参考种,在种子发育的不同阶段共鉴定出5 104个DEG,3个时期均差异表达的基因有78个(图 7b)。在每个阶段,下调表达的基因远超过上调表达的基因,分别是1 420∶787、566∶294、3 240∶785(表 5),分布于B、C染色体组的差异表达基因均表现为下调表达的基因数目显著多于上调表达基因。
-
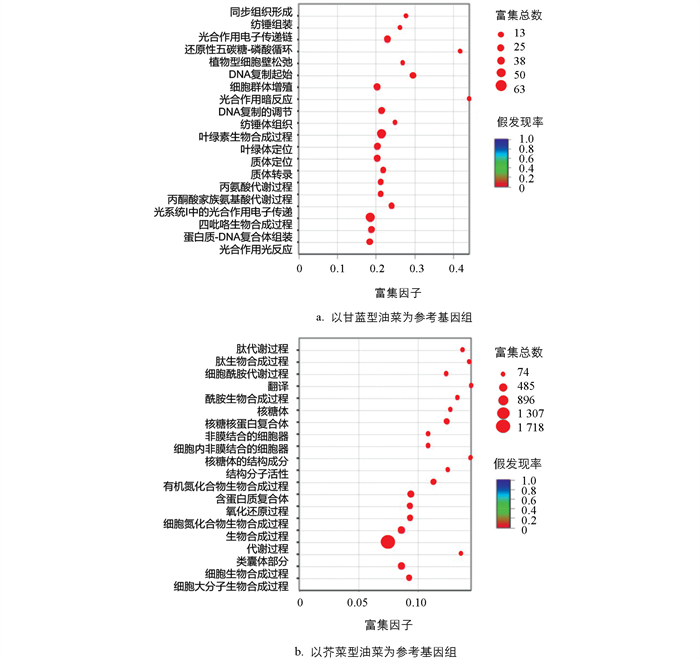
以甘蓝型油菜为参考种共得到5 495个差异表达基因,GO富集分析结果显示,20个GO terms被显著富集(p值<0.05),但并未检测到硫苷代谢相关的GO terms被显著富集(图 8a);对所有740个GO terms分析发现,其中GO:0019760被富集到参与硫苷代谢,共包含62个差异表达基因。以芥菜型油菜为参考种共得到5 104个差异表达基因,进一步GO富集分析,结果显示,共有20个GO terms被显著富集(p值<0.05),其中GO:0008152富集了1 718个差异表达基因,参与代谢过程(图 8b),但未检测到硫苷代谢相关的GO terms被显著富集。
-
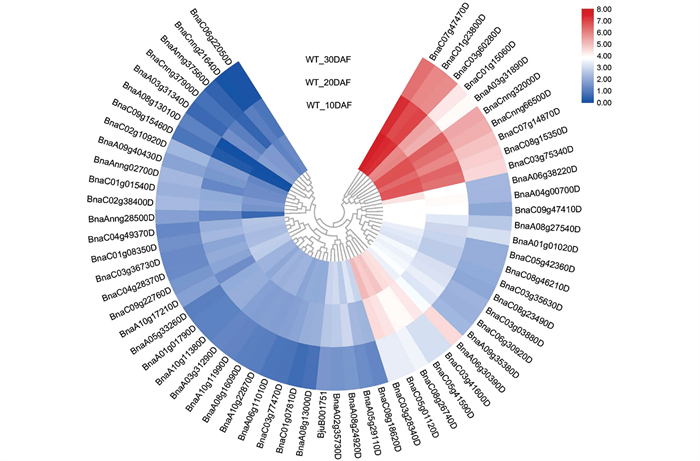
进一步对GO富集分析得到的埃塞俄比亚芥菜硫代葡萄糖苷合成路径中的差异表达基因进行聚类热图分析(图 9),包括以甘蓝型油菜为参考种得到的62个差异表达基因及以芥菜型油菜为参考种得到的1个差异表达基因,聚类结果显示参与硫代葡萄糖苷合成相关的63个差异表达基因聚为3个亚类,即随种子成熟显著上调表达的差异表达基因、显著下调表达的差异表达基因以及差异不显著的部分差异表达基因。其中仅有10个差异表达基因随种子成熟呈显著上调表达,分别为BnaC07g47470D、BnaC01g23800D、BnaC03g60280D、BnaC01g15060D、BnaA03g31890D、BnaCnng32000D、BnaCnng66500D、BnaC07g14870D、BnaC08g15350D、BnaC03g75340D。
2.1. BcMYB28的克隆及序列分析
2.2. CRISPR/Cas9载体构建及埃塞俄比亚芥菜的遗传转化
2.3. UHPLC-MS代谢物的测定及差异代谢物筛选
2.4. 转录组测序分析埃塞俄比亚芥菜硫苷代谢中的关键调节因子
2.5. 差异表达基因(DEGs)的检测及聚类分析
2.6. GO和KEGG富集分析
2.7. 埃塞俄比亚芥菜种子中参与硫代葡萄糖苷合成的DEGs聚类热图分析
-
MYB28基因在调控植物次生代谢途径中发挥着重要作用,特别是与硫代葡萄糖苷的合成密切相关[24-26]。本研究通过TA克隆及测序,获得了1个BcMYB28序列。将该序列提交至埃塞俄比亚芥菜数据库进行比对分析,鉴定出埃塞俄比亚芥菜中共有6个BcMYB28同源基因。这些基因序列与芸薹属作物中已知的MYB28基因序列的相似度为74.6%~95.8%,该发现揭示了MYB28基因在进化过程中的保守性。本研究通过CRISPR/Cas9成功构建了BcMYB28基因的双靶点敲除载体,并转化至埃塞俄比亚芥菜中,观察发现与埃塞俄比亚芥菜WT相比,T1代转化植株(CR)的叶片呈现出表面光亮、无蜡质层、无刺毛等显著表型变化。这些表型变化可能源于MYB28基因敲除后,其下游调控途径的紊乱。MYB28转录因子在植物体内扮演着重要的调控角色,它参与调控多种生物过程,包括芥子油苷代谢、蜡质合成以及抗病抗虫作用等。芥子油苷是十字花科植物中一类重要的次生代谢产物,具有抗病虫害和抗癌等生物活性。MYB28通过调控芥子油苷合成途径中的关键酶基因来影响芥子油苷的合成水平。当MYB28基因被敲除后,芥子油苷的合成将受到抑制[27],从而影响植物对病虫害的防御能力以及相关信号转导途径的正常运作。此外,MYB28还参与调控植物蜡质层的合成,蜡质层是植物表面的一层重要屏障,可以保护植物免受外界生物和非生物胁迫的侵害,MYB28通过调控蜡质合成基因的表达来影响蜡质层的形成。当MYB28基因被敲除后,蜡质层的合成将受到影响,导致植物对病虫害的防御能力降低[28]。在小麦中,MYB28还参与调控韧皮部防卫反应,这是植物抵御系统性侵染的病原微生物的一种有效机制[29]。当MYB28基因被敲除后,韧皮部防卫反应将受到影响,从而影响植物对病原微生物的抵御能力。
MYB28基因在硫代葡萄糖苷合成途径中起着关键调控作用,通过调控与硫代葡萄糖苷合成相关的酶基因(如侧链延长酶基因BCAT4、MAM1,中心结构合成酶基因CYP79F1等)的表达,来影响硫代葡萄糖苷的合成[30]。硫代葡萄糖苷与植物叶片表面蜡质层及刺毛的形成不受单个SAM细胞层的控制,而是受到细胞层间相互作用的影响[31]。因此硫代葡萄糖苷合成减少导致了SAM细胞层的相互作用,形成叶片表面光亮、无蜡质层、无刺毛等表型。此外,MYB28基因参与调控水杨酸(SA)和茉莉酸(JA)等激素的信号途径,来影响植物的防御反应和生长发育[32-33]。当MYB28基因被敲除后,这些信号途径可能受到干扰,从而导致植物生长发育的异常。
半胱氨酸是硫代葡萄糖苷合成中的关键前体物质,其通过一系列酶促反应转化为硫代葡萄糖苷的核心结构[16-17]。MYB28基因的敲除可能影响了这些酶促反应的进行,从而导致半胱氨酸向硫代葡萄糖苷的转化受阻。丝氨酸在硫代葡萄糖苷的侧链延长过程中具有重要作用[34],虽然丝氨酸本身并不直接参与硫代葡萄糖苷的合成,但其作为关键中间体的前体物质,对硫代葡萄糖苷的多样性和复杂性具有重要影响。本研究通过差异代谢物富集分析,找到甘氨酸、丝氨酸和苏氨酸代谢被显著富集,MYB28基因的敲除可能间接影响了丝氨酸的代谢途径,进而影响了硫代葡萄糖苷的侧链结构和含量。甘氨酸和苏氨酸的代谢变化可能通过影响植物体内的氨基酸平衡和能量代谢,间接影响硫代葡萄糖苷的合成和积累。
在对WT形成期种子进行转录组测序分析发现,埃塞俄比亚芥菜的清洁读段在比对到甘蓝型油菜和芥菜型油菜基因组时,比对率为51.06%~72.2%。埃塞俄比亚芥菜的染色体组构成主要源自B、C组染色体,通过其与甘蓝型油菜和芥菜型油菜的染色体比对,C01和C03在C染色体组中的比对率最高,而B05在B染色体中的比对率较高。芥菜型油菜的基因组被广泛用于与其他芸薹属植物的比较分析,尤其是与埃塞俄比亚芥的对比研究。这种比对率普遍较高,可能与两者在染色体组构成上的相似性有关[35]。由于埃塞俄比亚芥和芥菜型油菜在基因组结构上的相似性,尤其是B基因组的共性,使得在进行基因组比对时能够找到更多的同源区域,从而导致较高的比对率。通过差异表达基因的聚类热图分析找到10个硫苷合成相关差异表达基因,揭示了这些基因在埃塞俄比亚芥菜种子发育过程中调节硫苷代谢的潜在功能。
本研究采用CRISPR/Cas9技术成功建立了埃塞俄比亚芥菜的遗传转化体系,为后续的DEGs的功能验证奠定了基础。通过运用差异表达基因的聚类热图分析方法,鉴定出与硫苷合成相关的差异表达基因,揭示了这些基因在埃塞俄比亚芥菜种子发育过程中的潜在作用。这些研究成果进一步加深了对埃塞俄比亚芥菜硫苷代谢途径的了解,然而,探索这些DEGs在改良埃塞俄比亚芥菜、提高抗逆性等方面的应用潜力,尚需进一步深入研究以阐明。
 DownLoad:
DownLoad: