





-
开放科学(资源服务)标识码(OSID):

-
冬虫夏草(Ophiocordyceps sinensis,以下简称虫草)是名贵中药材,具有独特的药用价值,含多种活性成分,如核苷、多糖、甾醇、蛋白质及氨基酸等,备受消费者的青睐[1-3]。由于过度采挖,天然冬虫夏草资源日渐匮乏,市场上的需求缺口越来越大[4]。随着室内人工培殖虫草技术的进步,能培殖并诱导出有子实体的虫草,但在外观品质和商品规格等级上,人工培殖的虫草与天然虫草仍存在差异,影响了人工培殖虫草的商品价值[5-6]。
现有研究表明,昆虫体色分为色素色和结构色[5-6],其中,黑色素是昆虫体色多样性形成过程中至关重要的色素。黑色素通过酚氧化酶激活系统合成,与先天免疫、硬化和色素沉积密切相关[7]。真菌颜色变化主要由褐变引起,如食药用菌双孢蘑菇、金针菇、猴头菌等,其中,酶促褐变起主导作用。酶促褐变形成的3个关键因素是底物(酚类物质)、酶和活性氧(ROS)[8-10]。张芳[11]的研究表明,双孢蘑菇发生褐变主要是通过影响活性氧代谢及酚类物质代谢进行调控。本课题组前期利用组织细胞学、转录组学、蛋白组学等对人工培育虫草进行了品质机制研究,发现虫草僵虫颜色变化与活性氧代谢、免疫耐受力代谢相关[12],精氨酸在虫草品质变化过程中起着重要作用[13]。龚佳惠等[14]研究发现,0.1 mmol/L精氨酸处理能够抑制绿芦笋丙二醛含量及木质素的生成,提高了抗氧化能力,维持了绿芦笋的品质;1 mmol/L精氨酸处理能够提高抗氧化系统活性,提高了石榴果实的抗冷性,延缓了果实冷害症状的发生[15]。用不同浓度的精氨酸处理白蘑菇研究发现,精氨酸能够影响白蘑菇的贮藏品质[16]。精氨酸酶参与了茉莉酸甲酯调控的双孢蘑菇采后子实体的品质维持[17]。对番茄不同成熟期果实内源精氨酸代谢的研究发现,精氨酸在果实成熟过程中具有独特的生理作用[18]。大量研究表明,精氨酸对改善品质具有重要作用。
本研究通过精氨酸处理不同发育时期的虫草,利用苏木精—伊红染色法(HE)结合光学显微镜观察组织细胞学变化,采用试剂盒检测不同发育时期样本的丙二醛(MDA)、过氧化氢(H2O2)含量以及多酚氧化酶(PPO)、过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)的活性变化;通过Illumina测序平台对3个时期的冬虫夏草进行转录组测序分析,挖掘与颜色变化相关的关键基因,并从活性氧代谢角度分析精氨酸在调控虫草品质中的功能,旨在为进一步解析虫草虫体颜色变化的成因奠定分子学基础。
HTML
-
5龄感菌虫(MC)、僵虫(ST)、子座长度0~1 cm冬虫夏草(FB),由重庆市中药研究院冬虫夏草研究所实验室人工培殖提供,饲养温度为(8±1) ℃,湿度为50%±5%,无光照。选取虫体大小、颜色、发育时期均一致的样本,采用10 mmol/L精氨酸溶液[16]浸泡处理MC、ST、FB 30 min,处理后5龄感菌虫(TMC)、僵虫(TST)与MC、ST一起培养1个月,子座长度0~1 cm冬虫夏草(TFB)与FB培养至采收期。常规人工培殖冬虫夏草在人工模拟天然冬虫夏草产区环境进行培殖。
-
取不同发育时期样本[5龄感菌虫(MC、TMC)、僵虫(ST、TST)、子座长度0~1 cm冬虫夏草(FB、TFB)]于2%戊二醛固定液中固定24 h,经梯度酒精(70%、75%、85%、90%和100%酒精各30 min)脱水,再放入酒精与二甲苯混合液(体积比1∶1)中浸泡15 min,随后放入二甲苯中浸泡30 min。待样品完全透明后,将其放入二甲苯与石蜡混合液中浸泡10 min,最后放入软蜡和硬蜡中各30 min,冷却凝固。将石蜡包埋的样品经石蜡切片机进行切片,每个组织切片厚度为5~7 μm,放于载玻片制片,37 ℃过夜烤片,HE染色,树胶封片,光学显微镜下观察样品的组织细胞学结构并拍照。
-
将-80 ℃保存的待测组织置于已用液氮预冷过的研钵中,加入液氮研磨成粉,准确称取0.1 g粉末,加入900 μL PBS缓冲液进行匀浆,8 500 r/min 4 ℃离心10 min,取上清液用于酶活性及相关含量分析,置冰上待测。
-
多酚氧化酶(PPO)试剂盒(微量法,PPO-1-Y)、超氧化物歧化酶(SOD)试剂盒(NBT法,SOD-1-Y)、过氧化氢酶(CAT)试剂盒(钼酸铵比色法,CAT-1-W)、过氧化物酶(POD)试剂盒(微量法,POD-1-Y)、丙二醛(MDA)试剂盒(微量法,MDA-1-Y)及过氧化氢(H2O2)试剂盒(微量法,H2O2-1-Y),参照说明书进行检测,由苏州科铭生物技术有限公司提供。
-
Total RNA提取采用mirVanaTM miRNA Isolation Kit试剂盒(Ambion,美国),检测RNA样品的纯度、浓度和完整性,按照TruSeq Stranded mRNA LT Sample Prep Kit试剂盒操作说明书进行cDNA文库构建,使用Aglient 2100 Bioanalyzer生物分析仪进行质检,质检合格后于Illumina HiSeqTM 2500高通量测序平台对文库进行测序。
-
转录组测序后产生的原始数据(Raw reads)使用Trimmomatic软件进行质控并去除接头,在此基础上过滤掉低质量碱基以及N碱基,最终得到高质量的Clean reads。将Clean reads使用hisat2比对到物种的参考基因组,冬虫夏草菌参考基因组来源于NCBI数据库(
https://www.ncbi.nlm.nih.gov/assembly/GCA_000448365.1/ ),软件参数为默认参数,通过基因组比对率评估样本的情况。 -
对得到的Unigene进行基因功能注释,利用GO(基因本位论数据库,
http://geneontology.org/ )和KEGG(京都基因与基因组百科全书数据库,http://www.kegg.jp/ )进行GO分类和注释、KEGG代谢通路分析,进而获得所有Unigene的功能注释信息。 -
利用DESeq软件对差异表达基因进行分析,结合Benjamini-Hochberg方法对原有假设检验得到的显著性p值进行校正,p<0.05为差异表达基因筛选的关键指标。以错误发现率FDR<0.05且|log2FC|>1(Fold Change,FC)作为筛选标准,并对筛选到的差异表达基因进行GO富集及KEGG通路富集分析。
-
为了验证转录组数据的可靠性,采用真菌18 S核糖体RNA作为内参基因,随机选取6个差异表达基因进行qRT-PCR验证,设计qRT-PCR特异引物(表 1),利用反转录试剂盒HiScript® Ⅱ 1st Strand cDNA Synthesis Kit(Vazyme,中国)将RNA反转录得到cDNA,采用SYBR Green Master Mix荧光定量试剂盒(Vazyme,中国)在荧光定量PCR仪上进行定量检测,每个样品3个重复,采用2-ΔΔCt法计算差异基因的相对表达量[19]。
-
采用Excel 2020进行数据整理,运用GraphPad Prism软件对数据进行显著性分析与作图。统计分析采用t检验,数据以 x±s表示。
1.1. 供试材料
1.2. 试验方法
1.2.1. 组织细胞学鉴定
1.2.2. 待测液提取
1.2.3. 活性氧代谢、MDA及H2O2含量测定
1.2.4. Total RNA提取与高通量测序
1.2.5. 序列筛选与组装
1.2.6. Unigene功能注释和分类
1.2.7. 差异表达基因的筛选
1.2.8. qRT-PCR分析
1.3. 数据分析
-
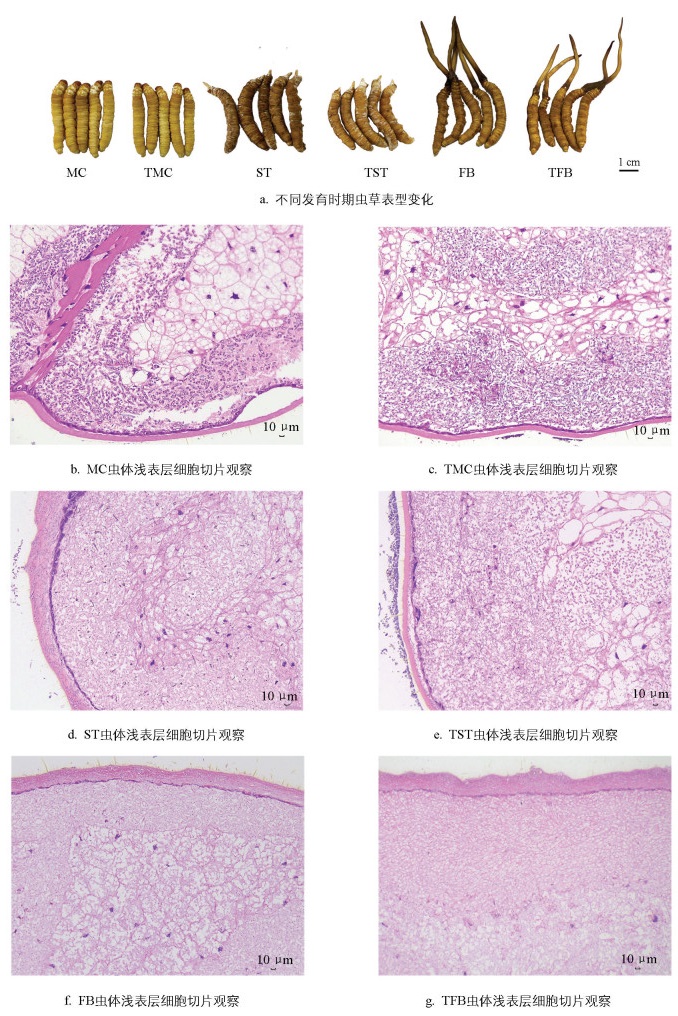
为明确精氨酸在虫草发育过程中如何影响品质,本研究对不同发育时期虫草进行虫体表型鉴定和组织细胞学分析。图 1a显示,与MC和FB相比,TMC和TFB虫体颜色无明显变化;与ST相比,TST虫体颜色更优。对各组织样本纵切,经HE染色后发现,与MC相比,TMC的组织细胞密度变小、组织结构更加紧凑(图 1b、1c);与ST相比,TST的组织细胞密度也变小、组织结构也更加紧凑(图 1d、1e)。虫草生长后期,与FB相比,TFB无明显变化(图 1f、1g)。由此表明,虫草生长发育前期易受精氨酸胁迫,影响了虫体浅表层的细胞结构。
-
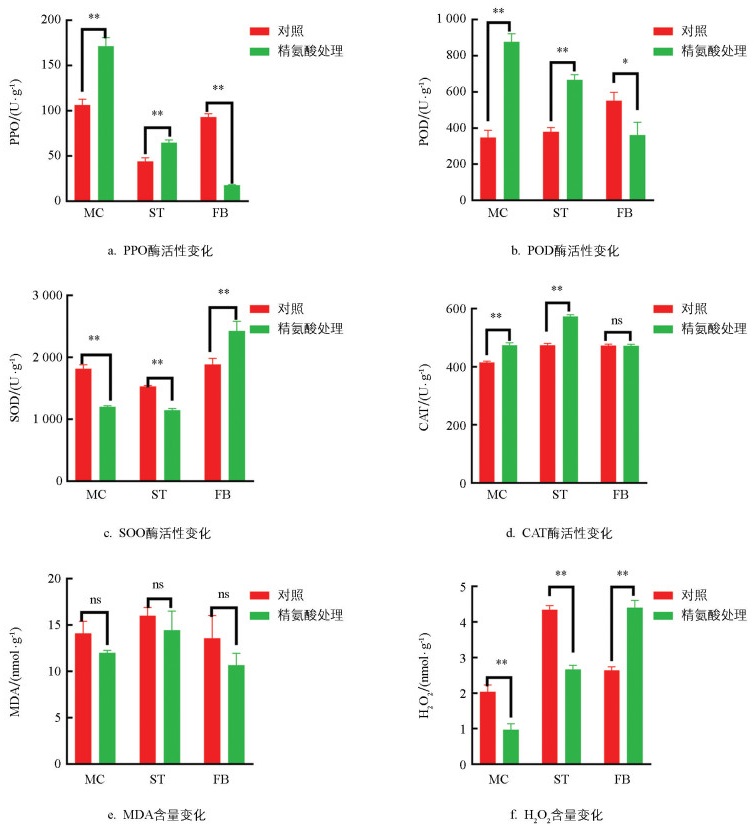
对各组织样本体内活性氧代谢相关酶(PPO、SOD、CAT、POD)活性、MDA和H2O2含量进行检测,结果表明(图 2),在虫草发育前期,与对照相比,TMC和TST的PPO、POD、CAT活性显著增加(p<0.01),SOD酶活性显著降低(p<0.01);在发育后期,与对照相比,TFB的PPO(p<0.01)、POD(p<0.05)活性显著降低,SOD活性显著增加(p<0.01),CAT活性无明显变化。整个发育周期中,精氨酸处理前后MDA含量较对照有所减少,但变化不明显(图 2e)。与对照相比,TMC和TST的H2O2含量显著降低(p<0.01),TFB的H2O2含量显著增加(p<0.01,图 2f)。综合表明,精氨酸处理不同发育时期的虫草后,活性氧代谢相关酶(PPO、SOD、POD、CAT)活性和H2O2含量变化差异有统计学意义。
-
为探究精氨酸调控虫草品质的关键基因,本研究利用RNA-seq技术对虫草3个发育时期(MC:5龄感菌虫;TMC:精氨酸处理的5龄感菌虫。ST:僵虫;TST:精氨酸处理的僵虫。FB:子座长度0~1 cm冬虫夏草;TFB:精氨酸处理的子座长度0~1 cm冬虫夏草)样本进行了分析,如表 2。测序总计得到8.944 2×108 Raw reads,经质量筛选后,得到8.636 9×108 Clean reads,共计1.253 5×1011 Clean bases,各样本的有效数据量为(6.88~7.07)×109,Q30碱基为95.15%~96.61%,平均GC为54.50%(表 2)。从测序产出数据质量评估结果可知,本次测序数据饱和度较高,可以满足后期的数据分析。
-
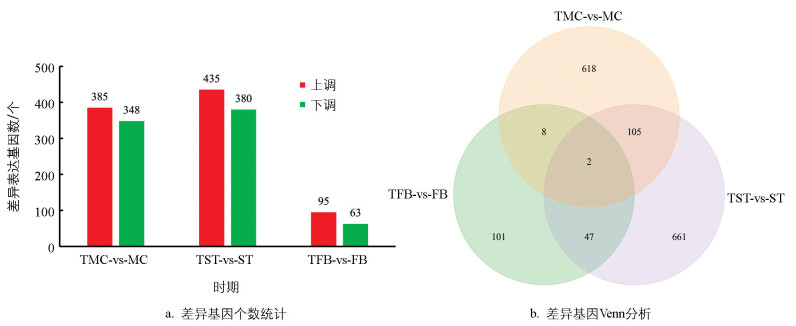
差异表达基因筛选结果如图 3,由柱状图、差异表达韦恩(Venn)图展示差异基因数目,TMC-vs-MC共有733个基因表达量发生了显著变化,其中上调表达的基因有385个,下调表达的基因有348个;TST-vs-ST共有815个基因表达量发生了显著变化,其中上调表达的基因有435个,下调表达的基因有380个;TFB-vs-FB共有158个基因表达量发生了显著变化,其中上调表达的基因有95个,下调表达的基因有63个。在这些基因中,总共有162个基因持续地发生显著变化。由此表明,采用精氨酸处理前期虫草,对其生长发育的影响较大;后期采用精氨酸处理,差异表达基因数量显著减少,表明精氨酸对虫草生长发育后期影响较小。
-
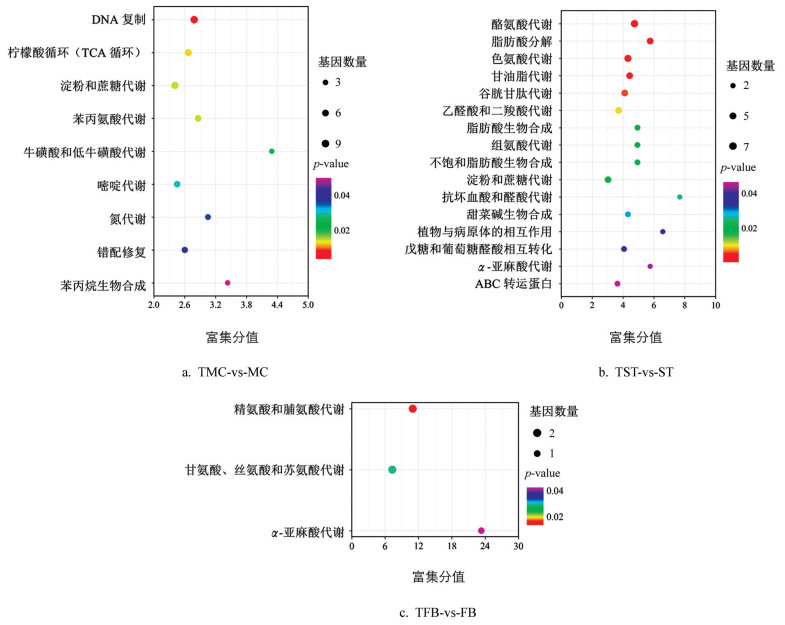
KEGG富集分析发现(图 4),这些差异表达基因主要富集在酪氨酸代谢通路(Tyrosine metabolism,ko00350)、精氨酸和脯氨酸代谢通路(Arginine and proline metabolism,ko00330)、DNA复制(DNA replication,ko03030)、色氨酸代谢通路(Tryptophan metabolism,ko00380)、苯丙氨酸代谢通路(Phenylalanine metabolism,ko00360)、苯丙烷生物合成通路(Phenylpropanoid biosynthesis,ko00940)上。
-
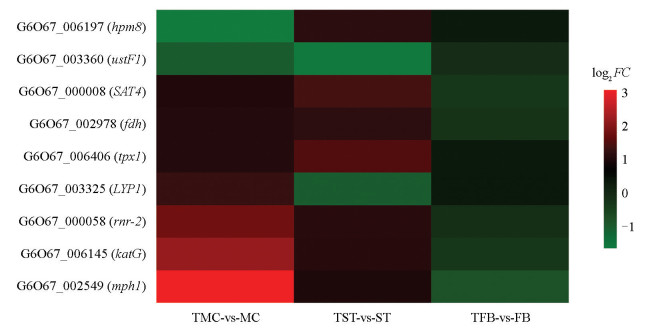
昆虫和食药用菌颜色变化主要由过氧化物酶、酚氧化酶、活性氧、黑化反应等调控[20-22]。由图 5可知,丝/苏氨酸蛋白激酶G6O67_002549(Serine/threonine-protein kinase,mph1),过氧化氢酶G6O67_006145(Catalase-peroxidase,katG),核糖核苷二磷酸还原酶G6O67_000058(Ribonucleoside-diphosphate reductase,rnr-2),丝/苏氨酸蛋白激酶G6O67_000008(Serine/threonine-protein kinase,SAT4),甲酸脱氢酶G6O67_002978(Formate dehydrogenase,fdh)在TMC-vs-MC和TST-vs-ST中表达显著上调,在TFB-vs-FB中表达下调;在整个发育周期G6O67_003360(ustF1)均表达下调。其中,有3个差异表达基因能显著富集到KEGG通路中,分别是过氧化氢酶katG、核糖核苷二磷酸还原酶rnr-2、甲酸脱氢酶fdh。值得关注的是,过氧化氢酶katG表达变化趋势与CAT酶活性变化相一致。
-
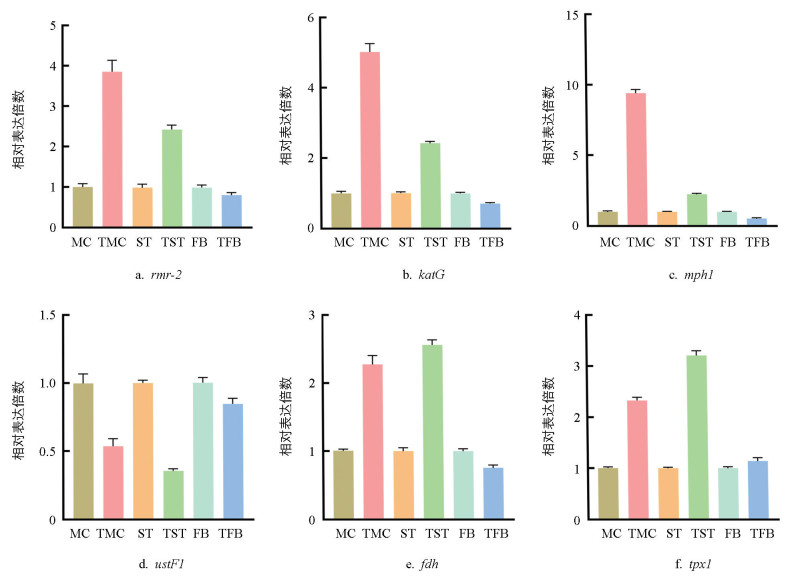
为了验证转录组数据的准确性,随机挑选6个差异表达基因进行qRT-PCR验证,结果显示这6个基因表达水平与转录组数据的变化趋势相一致(图 6),由此表明,转录组测序数据可靠。
2.1. 不同发育时期组织细胞学分析
2.2. 不同发育时期虫草活性氧代谢相关酶活性、MDA及H2O2含量变化
2.3. 转录组测序质量分析
2.4. 差异表达基因分析
2.5. 差异表达基因KEGG富集分析
2.6. 冬虫夏草品质调控关键基因挖掘
2.7. qRT-PCR验证
-
为明确人工培殖的虫草颜色变化品质机制,本研究对精氨酸处理不同发育时期虫草的表型及细胞学特征进行了分析。结果表明,精氨酸通过调控活性氧代谢相关酶活性(尤其是过氧化氢酶),进而影响虫草虫体颜色的变化。经查阅文献资料发现,无论是探究昆虫颜色变化,还是探究食药用菌颜色变化涉及的酶促褐变,都与活性氧代谢密切相关[22-25]。
差异表达基因筛选分析发现,TMC-vs-MC共有733个基因、TST-vs-ST共有815个、TFB-vs-FB共有158个基因表达量发生了显著变化,其中,162个基因持续发生显著变化。这些差异表达基因主要富集在酪氨酸代谢通路(ko00350)、精氨酸和脯氨酸代谢通路(ko00330)、DNA复制(ko03030)、色氨酸代谢通路(ko00380)、苯丙氨酸代谢通路(ko00360)、苯丙烷生物合成通路(ko00940)上。研究结果表明,酪氨酸代谢通路、苯丙氨酸代谢通路、精氨酸和脯氨酸代谢通路、苯丙烷生物合成通路相关基因可能参与了虫草发育过程中虫体颜色的变化,这与前人报道相一致。例如,酪氨酸酶是黑色、白色、黄色3种色素前体物质合成过程中的关键限速酶,酪氨酸酶活性或其基因表达受影响时,会造成底物大量积累并沿着某一色素合成方向进行反应[26-27]。鳞翅目昆虫家蚕已有多个与黑色素合成信号通路、苯丙氨酸代谢通路、苯丙烷生物合成通路相关功能基因被报道,如TH(酪氨酸羟化酶)[28]、DDC(多巴脱羧酶)[29]、yellow/laccase2(漆酶)[30]等。此外,精氨酸不仅能保持蓝莓的品质,还能激活其抗氧化系统[31]。精氨酸能有效抑制柑橘的酶促褐变[32],还能抑制刚切开的苹果和生菜的褐变[33]。本研究中筛选到1个过氧化氢酶基因katG,该基因能显著富集到苯丙氨酸代谢通路、苯丙烷生物合成通路上,这些代谢通路都与褐变、活性氧、黑化等过程密切相关[22-25]。由此表明,精氨酸处理虫草后,通过调控过氧化氢酶活性或过氧化氢酶基因的表达,进而激活抗氧化系统,调节虫草颜色的变化。
-
精氨酸处理不同发育时期的虫草,活性氧代谢相关酶(PPO、SOD、POD、CAT)活性和H2O2含量变化差异有统计学意义。在虫草发育过程中,TMC-vs-MC总共有733个、TST-vs-ST共有815个、TFB-vs-FB共有158个基因表达量发生了显著变化,其中,162个基因持续发生显著变化。这些差异表达基因主要富集在酪氨酸代谢通路、苯丙氨酸代谢通路、精氨酸和脯氨酸代谢通路、色氨酸代谢通路、DNA复制、苯丙烷生物合成通路上。过氧化氢酶基因katG表达变化趋势与CAT酶活性变化相一致。
 DownLoad:
DownLoad: