





-
开放科学(资源服务)标识码(OSID):

-
转录因子(Transcription Factor,TF)是一类通过与特定顺式作用元件结合来调控基因表达的蛋白质,在植物应对非生物胁迫、激素信号转导以及生长发育过程中发挥着重要作用[1-2]。已知NAC(NAM、ATAF1/2和CUC2)结构域蛋白是在多种植物发育过程中具有不同功能的植物特异性转录因子。首个NAC转录因子是从矮牵牛(Petunia hybrida)中发现的NAM(No Apical Meristem)基因,它与矮牵牛胚胎的生长和发育密切相关[3],因携带NAM突变的矮牵牛胚胎无法发育出顶端分生组织,导致幼苗缺乏根和叶[4]。随后,在拟南芥(Arabidopsis thaliana (L.) Heynh.)中也发现了两个相关的NAC转录因子ATAF1/2和CUC2[5-6],NAC转录因子的名称由“NAM” “ATAF” “CUC”3个基因的首字母组成。NAC转录因子由N末端的靶结合域和与其他蛋白质相互作用的高度保守的C末端域组成。N末端的保守结构域最初在4个基因(NAM、ATAF1、ATAF2和CUC2)中被发现,随后进一步将其划分为5个亚结构域(A至E)。其中,A、C和D结构域在不同物种的NAC成员中高度保守,而B和E结构域则表现出更多的多样性[2]。
NAC蛋白作为植物特异性转录调节因子中最大的一类,目前已在水稻(Oryza sativa L.)[7]、玉米(Zea mays L.)[8]、马铃薯(Solanum tuberosum L.)[9]等多种植物中得到鉴定。研究表明,NAC蛋白在植物应对各种逆境胁迫时发挥着独特的功能,能够直接或间接调控转录因子下游的应激相关基因,或通过与其他蛋白质相互作用,使植物响应外界环境的各种胁迫。例如,大豆(Glycine max (L.) Merr.)中与拟南芥ATAF1同源的GmNAC109在盐胁迫下能够显著诱导其表达[10];花椒(Zanthoxylum bungeanum Maxim.)的多个NAC转录因子(如ZbNAC7/18/47/72/79)在抗旱过程中起着关键作用[11]。此外,NAC转录因子还会在冷胁迫下受到诱导,参与植物对低温胁迫的反应[12]。番茄(Solanum lycopersicum L.)的SlNAM1和SlNAC1在低温胁迫下表达上调,有助于提高转基因植物的抗寒能力[13-14];茶树(Camellia sinensis (L.) O. Kuntze)的CsNAC17/26/30在低温胁迫下的表达逐渐增加,其中CsNAC17和CsNAC26的诱导作用尤为显著,常用作茶树的抗冻性分子标记进行性状鉴定[15]。
NAC转录因子除参与植物应激反应外,还广泛参与根系生长、花器官发育、种子萌发、植物衰老和诱导细胞死亡等生物学过程[16-19]。水稻OsNAC2通过调控赤霉素(Gibberellic Acid,GA)途径影响植株高度和开花时间,OsNAC54能够直接调节CGGs基因活性并促进ABA INSENSITIVE 5(ABI5)的表达,从而间接影响叶片衰老[20-21];拟南芥AtNAC1通过抑制细胞分裂、DNA修复和细胞分化的关键调节因子E2Fa的转录,来维持根分生组织的功能和促进根生长[22];芜菁(Brassica campestris L.) BcNAC2经组织原位杂交后在胚囊中高度表达,说明NAC转录因子与种子或胚胎发育有关[23]。以上研究表明,NAC转录因子可通过外界胁迫或植物信号调控其表达,进而参与多种生物学过程。
蒺藜苜蓿(Medicago truncatula)是豆科(Leguminosae)苜蓿属(Medicago L.)的一年生植物,因其具有基因组小、自花授粉、生长周期短和遗传转化效率高等优点,被广泛用作研究豆科植物的模式植物。目前,蒺藜苜蓿的全基因组数据已经公布,这为深入研究其基因调控、挖掘潜在遗传信息创造了有利条件[24]。对蒺藜苜蓿的研究不仅有助于理解豆科植物在生长发育过程、激素调控、信号转导及逆境胁迫响应等方面的理论,还具有重要的应用价值。NAC转录因子尽管在拟南芥和水稻等模式植物中已有相关研究,但在蒺藜苜蓿中的研究相对较少。本研究基于课题组前期的蛋白互作筛选,在蒺藜苜蓿MtUFO上游调控因子中鉴定出MtNAC47基因,克隆了蒺藜苜蓿MtNAC47基因,并进行了生物信息学分析、亚细胞定位、转录自激活检测及表达特征分析,旨在为该基因功能的进一步研究奠定理论基础。
HTML
-
试验材料为蒺藜苜蓿野生型(M. truncatula ‘R108’),由北京林业大学草业与草原学院分子实验室提供。试验耗材RNA提取试剂盒(CW0598S)购于北京康为世纪生物公司,2× RapidTaq Master mix(P222)购于南京诺唯赞公司,OMEGA PCR纯化试剂盒(D6492)购于广州飞扬生物工程有限公司,大肠杆菌感受态细胞DH5α(KTSM101L)购于北京康体生命公司,反转录试剂盒(AG11728)、qRT-PCR试剂盒(AG11733)购自湖南艾科瑞生物工程有限公司,Minimal SD Base(PM2030)、DO Supplement-Trp(PM2250)、X-α-gal(CX11922)、金担子素(Aureobasidin A)(CA2332)购自北京酷来搏科技有限公司,农杆菌感受态EHA105、酵母菌株Y2HGold、酵母诱饵表达载体pGBKT7及植物表达载体3302Y均为实验室保存。
-
使用Primer Premier 5.0软件进行引物设计(表 1),MtNAC47-F/R用于克隆MtNAC47基因;3302Y-MtNAC47-F/R用于构建亚细胞定位植物表达载体;pGBKT7-MtNAC47-F/R用于构建酵母表达载体;T7 promoter和3′BD用于检测酵母表达载体;MtNAC47-RT-F/R用于MtNAC47基因的表达分析,MtActin-F/R用于内参基因MtActin的表达分析[25]。
-
以蒺藜苜蓿‘R108’为植物材料,使用RNA提取试剂盒提取植物总RNA,随后反转录为cDNA。以MtNAC47-F/R为引物,蒺藜苜蓿cDNA为模板,利用2× RapidTaq Master mix进行PCR扩增,反应体系为20 μL。反应程序为:95 ℃ 3 min,95 ℃ 15 s,55 ℃ 20 s,72 ℃ 15 s,34个循环,72 ℃延伸5 min,12 ℃保温。反应结束后利用琼脂糖凝胶对PCR产物进行电泳检测,并借助PCR纯化试剂盒回收目的条带连接至克隆载体pMD19-T。使用大肠杆菌感受态细胞DH5α对连接产物进行转化,吸取适量菌液涂布在含有氨苄青霉素(100 mg/L)的固体LB培养基上,倒置于37 ℃培养箱中12 h。挑取单克隆菌落进行PCR检测,将符合目的基因片段大小的菌液送至北京睿博兴科生物公司测序,将测序正确的质粒命名为pMD-MtNAC47。
-
在NCBI网站对MtNAC47蛋白序列进行Blast比对,并下载同源蛋白序列。通过MEGA7.0软件的邻接法(neighbor-joining,NJ)构建系统发育树,Bootstrap重抽样次数默认为1 000次,以评估系统发育树的可靠性。利用DNAMAN 9.0对MtNAC47及其他物种同源蛋白序列进行多序列比对分析。根据SOPMA(
https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html )和SWISS-MODEL(https://swissmodel.expasy.org/interactive/QdTzAC/models/ )网站预测蛋白质的二级和三级结构。使用ExPASy-ProtParam(https://web.expasy.org/protparam/ )分析MtNAC47蛋白的理化性质,使用SignalP 4.1(https://services.healthtech.dtu.dk/services/SignalP-4.1 )预测信号肽。在TMHMM网站(https://services.healthtech.dtu.dk/service.php TMHMM-2.0)预测MtNAC47的跨膜结构域,并通过Expasy网站(https://www.expasy.org/ )分析其亲疏水性。借助NetPhos(https://services.healthtech.dtu.dk/service.php NetPhos-3.1)进行磷酸化位点预测。从蒺藜苜蓿基因组数据库中选择MtNAC47上游的2 000 bp作为启动子序列,通过PlantCARE(https://bioinformatics.psb.ugent.be/webtools/plantcare/ )进行启动子分析,并用TBtools进行数据可视化。从NCBI下载13个同源蛋白物种的基因组文件,分析NAC基因家族的基因结构,并在NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )获取NAC结构域可视化分析结果。使用MEME(https://meme-suite.org/meme/tools/meme )分析NAC蛋白的保守基序,最后利用TBtools的Gene Structure View进行基因结构、保守基序和结构域的可视化。 -
从测序正确的菌液中提取质粒,使用特异性引物3302Y-MtNAC47-F/R,以pMD-MtNAC47作为模板进行PCR扩增。经琼脂糖凝胶电泳鉴别后,回收纯化符合预期条带的PCR产物。用限制性内切酶Nco Ⅰ对表达载体3302Y进行酶切,随后采用无缝克隆技术按1∶1比例连接纯化后的PCR产物及酶切产物,转化至大肠杆菌感受态细胞DH5α中,吸取并将菌液涂布在LB固体培养基(含50 mg/L卡那霉素)上,37 ℃培养箱中倒置过夜培养后挑取单菌落进行PCR检测并测序验证,测序正确的质粒命名为3302Y-MtNAC47。将质粒3302Y-MtNAC47转入农杆菌感受态EHA105中,进行农杆菌菌液摇菌,在培养箱中避光震荡2~3 d,直至OD 600为0.6~0.8时,将重悬菌液注射到1月左右生长良好的烟草叶片细胞下表皮中,避光培养48 h,之后借助激光共聚焦显微镜(SP8)观察MtNAC47在细胞中的位置。
-
以pMD-MtNAC47质粒为模板,BD-MtNAC47-F/R为引物,进行PCR扩增,同时使用限制性内切酶EcoR I对酵母表达质粒pGBKT7进行单酶切。将PCR产物和酶切后的质粒分别进行纯化,随后连接重组。连接产物通过大肠杆菌感受态转化、卡那霉素筛选及菌落PCR检测后,将含有目标条带的菌液送至生物公司进行测序分析。测序正确的质粒命名为BD-MtNAC47,提取质粒并利用PEG/LiAc转化质粒BD-MtNAC47至Y2HGold酵母感受态细胞中,吸取100~200 μL菌液均匀涂布在SD/-Trp固体培养基上,于30 ℃的酵母培养箱中培养2~3 d,挑取单菌落进行PCR验证。
-
将含有BD-MtNAC47的酵母菌液使用0.9% NaCl稀释至10倍、100倍、1 000倍后,分别滴加在SD/-Trp,SD/-Trp/X-α-Gal/AbA固体培养基上。在酵母培养箱中倒置培养3~5 d。培养结束后观察酵母菌生长情况并进行拍照记录。
-
挑选健康饱满的蒺藜苜蓿种子,用砂纸轻微打磨种皮,使其破损。将种子置于有吸水滤纸的培养皿中,在4 ℃下进行春化处理3~4 d。随后,将种子放入光照16 h/25 ℃、黑暗8 h/23 ℃的培养箱中,直至根系发育。接着将其移栽到花盆中,并在光周期为光照16 h、黑暗8 h,明暗温度分别为25 ℃、23 ℃,湿度为56%的人工气候箱中继续培养。选取3月龄、长势良好一致的蒺藜苜蓿,分别取其根、茎、叶、花和果5部分组织,同时分别对单株的全部叶片喷施50 μmol/L的脱落酸(Abscisic Acid,ABA)、10 μmol/L的茉莉酸甲酯(Methyl Jasmonate,MeJA)、10 μmol/L的水杨酸(Slicylic Acid,SA)、10 μmol/L的GA进行激素诱导,并将单株植株置于4 ℃低温环境中进行胁迫处理。在0 h、0.5 h、1 h、3 h、6 h、9 h、12 h 7个时间点对叶片组织进行取样,每个处理进行3次生物学重复,样品用液氮速冻后放在超低温冰箱-80 ℃保存。对样品进行RNA提取并反转录获得cDNA,以MtActin作为内参基因,MtNAC47-RT-F/R为特异性引物,根据qRT-PCR试剂盒说明书使用荧光定量PCR仪器进行荧光定量,试验样品均采用3次生物学重复,所得数据采用2-ΔΔCt法,使用Excel 2019进行分析处理。
1.1. 材料
1.2. 方法
1.2.1. 引物设计
1.2.2. MtNAC47基因克隆
1.2.3. MtNAC47的生物信息学分析
1.2.4. 表达载体构建及亚细胞定位
1.2.5. BD-MtNAC47酵母表达载体构建及Y2HGold酵母感受态细胞的转化
1.2.6. 转录自激活作用检测
1.2.7. MtNAC47基因的表达模式分析
-
以MtNAC47-F和MtNAC47-R为特异性引物,蒺藜苜蓿cDNA为模板进行PCR扩增。反应结束后得到一条清晰且符合预期的条带,长度约为1 000 bp,随后送至生物公司进行测序验证(图 1a)。测序结果表明,MtNAC47长度为1 023 bp,条带长度与目的基因序列长度一致,测序结果与目标基因序列一致性比对结果为100%,证明成功克隆了蒺藜苜蓿MtNAC47基因。
-
对蒺藜苜蓿MtNAC47蛋白和红三叶(Trifolium pratense L.)、豌豆(Pisum sativum L.)、毛苕子(Vicia villosa Roth.)等40个物种中的NAC同源蛋白进行系统发育树构建。结果表明,MtNAC47与GbNAC47、RcNAC47、TpNAC47、DzNAC47等12个蛋白聚合为一个亚簇分支,和红三叶TpNAC47亲缘关系最近,推断二者具有相似的生物学作用(图 1b)。对蒺藜苜蓿MtNAC47基因的保守结构域进行预测,发现MtNAC47基因属于NAM超家族。对蒺藜苜蓿及其他12个物种的氨基酸序列进行多序列比对分析,结果显示蒺藜苜蓿MtNAC47蛋白与毒羊豆(Gastrolobium bilobum R.Br.)、蓖麻(Ricinus communis L.)、红三叶等同源蛋白相似度较高(图 1c)。
-
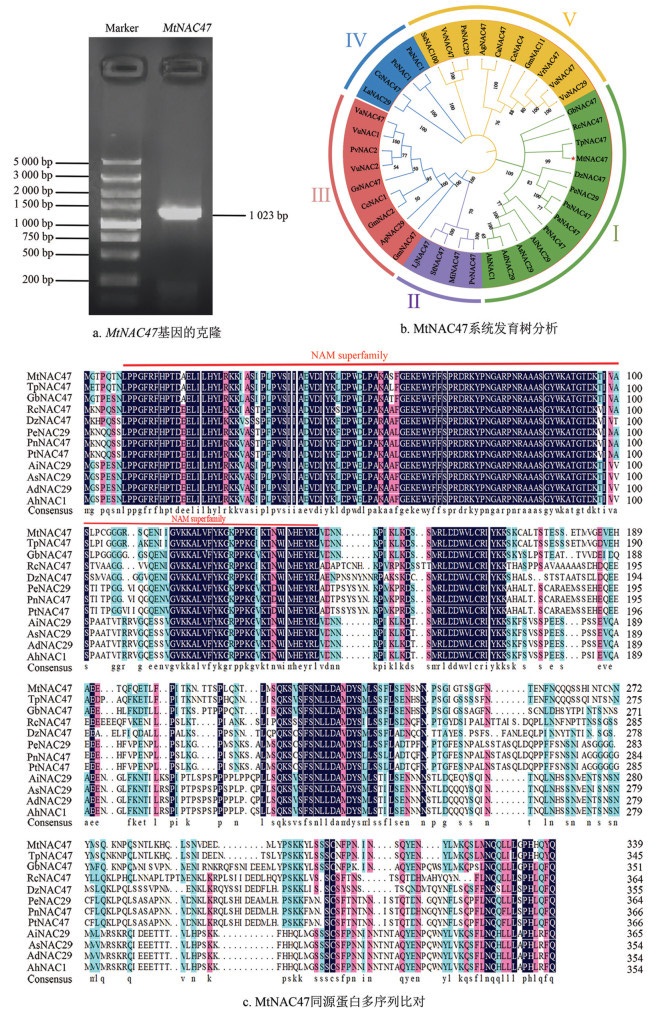
蛋白二、三级结构分析表明MtNAC47蛋白由4种二级结构模块组成,其中:无规则卷曲占比最高,为80%;延伸链为11.76%;α-螺旋占比为8.24%(图 2a、2b)。蛋白质跨膜结构分析表明MtNAC47是非跨膜性蛋白,信号肽预测显示MtNAC47蛋白无信号肽,为非分泌蛋白或非膜蛋白,意味着该蛋白不被转运到细胞外或跨越细胞膜(图 2c、2d)。对MtNAC47蛋白的亲疏水性进行分析,发现该蛋白含有多个亲水区和疏水区(图 2e)。磷酸化位点分析显示MtNAC47蛋白共具有43个磷酸化位点,包括26个丝氨酸磷酸化位点、11个苏氨酸磷酸化位点和6个酪氨酸磷酸化位点(图 2f)。
蛋白理化性质分析表明,MtNAC47编码340个氨基酸,分子式C1706H2652N462O521S15,丝氨酸占比最高,为10.9%,其次为亮氨酸,占比9.4%,蛋白分子量为38.45 kg/mol,理论pI值8.87,为碱性蛋白,不稳定指数为41.63,总平均亲水指数为-0.639,说明MtNAC47是一个亲水不稳定的碱性蛋白(表 2)。
-
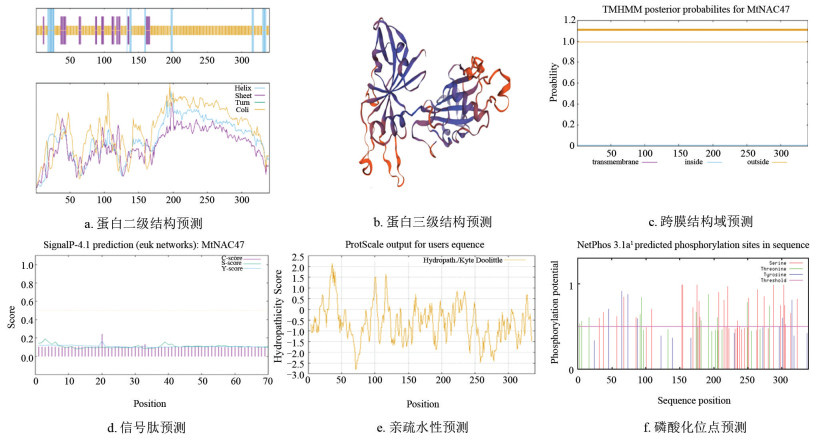
同源蛋白结构域分析表明MtNAC47蛋白与同源分支蛋白均包含NAM保守结构域,属于NAC转录因子家族(图 3a)。motif分析发现,该蛋白在13个物种中均包含多个保守基序(如motif1/2/3/4/5/6),且这些基序的位置差异极小(图 3b)。通过分析MtNAC47与同源基因的外显子和内含子结构,预测该基因可能编码的蛋白质结构和功能。基因结构分析表明除PaNAC47和TpNAC47外,其他聚类中的基因结构较为相似,在基因长度、外显子和内含子数量上差异较小(图 3c),蛋白保守基序分析显示氨基酸残基的变异幅度较小且数量较多(图 3d)。
-
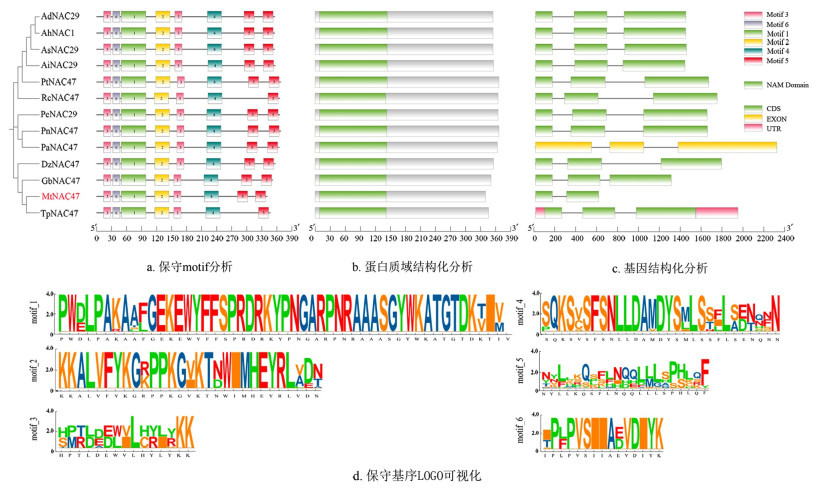
顺式作用元件分析发现MtNAC47上游的启动子区除含有光响应元件(Gap-box、GATA-motif等)外,还具有多种与植物激素及非生物胁迫相关的反应元件,如MeJA反应元件(CGTCA-motif、TGACG-motif),ABA反应元件(ABRE),GA反应元件(TATC-box)以及厌氧诱导(ARE)和低温响应(LTR)元件,说明MtNAC47基因在调控蒺藜苜蓿的生长发育、激素信号转导和非生物胁迫响应等多种生理过程中发挥重要作用(图 4a、4b)。
-
为检测MtNAC47蛋白在细胞中的定位情况,将携带GFP的3302Y-MtNAC47及3302Y空载的菌液分别注射到烟草下表皮细胞,暗培养48 h后观察荧光信号。在激光共聚焦显微镜下的观察结果表明,处理组烟草表皮细胞在细胞核和细胞质中检测到MtNAC47-GFP融合荧光蛋白,对照组烟草则在整个表皮细胞中均可检测到荧光信号,说明MtNAC47蛋白定位于细胞核和细胞质(图 5)。
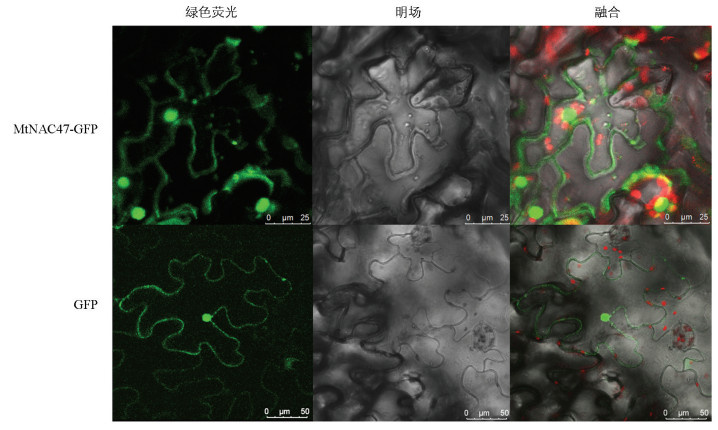
-
为研究MtNAC47是否具有转录自激活活性,将MtNAC47基因连接到酵母表达载体pGBKT7-BD上,转入酵母感受态Y2HGold中;并以BD-MsMYB33为阳性对照,空白载体BD-Lam为阴性对照,稀释后点涂到SD/-Trp和SD/-Trp/X-α-Gal/AbA固体培养基上,进行3~5 d的培养。结果显示,在SD/-Trp培养基上菌落正常生长;在SD/-Trp/X-α-Gal/AbA固体培养基上,菌落均能够正常生长且表现为蓝色,表明BD-MtNAC47存在转录自激活活性(图 6)。

-
为更好研究MtNAC47基因在蒺藜苜蓿器官发育中的作用,通过荧光定量技术测定MtNAC47在蒺藜苜蓿不同组织中的表达量。分析表明,MtNAC47在根、茎、叶、花和果中均有表达,在花中的相对表达量最高,其次是根和茎,在果中的相对表达量最低(图 7a)。对蒺藜苜蓿分别进行ABA、MeJA、SA、GA激素处理及低温胁迫处理,以分析MtNAC47基因在不同激素处理及非生物胁迫下的应答情况。结果显示,ABA处理后,MtNAC47的相对表达量呈先下降后上升的趋势,在9 h时达到顶峰,是0 h时的1.4倍(图 7b);MeJA处理后,MtNAC47的相对表达量显著下降,于9 h时达到低谷(图 7c);SA处理后,MtNAC47的相对表达量整体表现为先升后降再回升的趋势,于3 h时到达最高值,是对照组的3.8倍,随后下降并在9 h出现回升(图 7d);GA处理后,MtNAC47的相对表达量先上升后下降,在6 h时达到最大值,上调到0 h的2.4倍(图 7e);经低温胁迫处理后,MtNAC47的相对表达量整体表现出先下降后上升,在6~9 h出现回升,随后下降并于12 h时出现最低值(图 7f)。
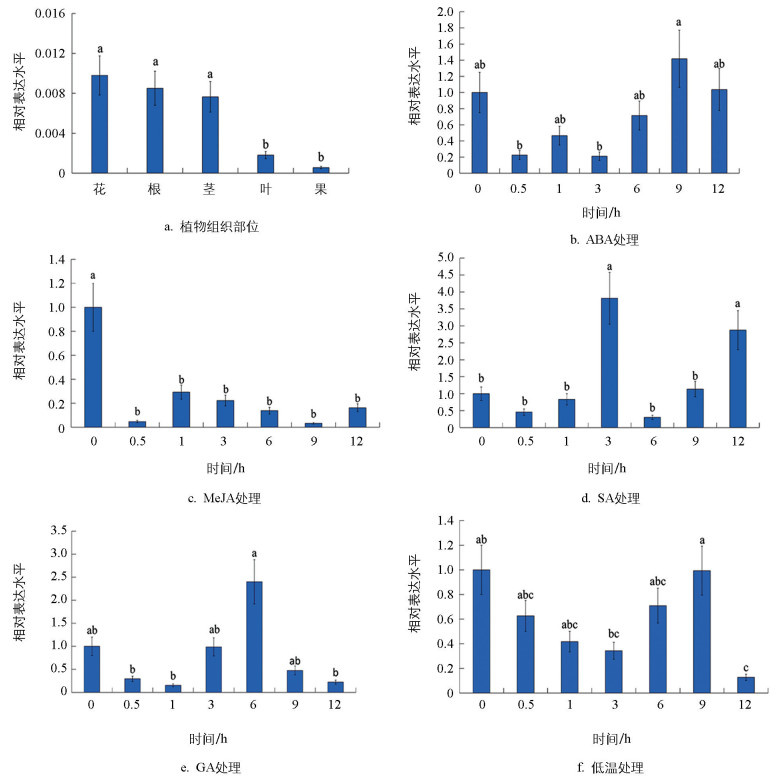
2.1. MtNAC47基因克隆
2.2. MtNAC47基因的生物信息学分析
2.2.1. MtNAC47系统发育树构建及同源蛋白比对
2.2.2. MtNAC47蛋白结构预测及理化性质分析
2.2.3. MtNAC47及其同源分支蛋白的保守基序和结构域分析
2.2.4. MtNAC47基因顺式作用元件分析
2.3. MtNAC47的亚细胞定位
2.4. 转录自激活检测
2.5. MtNAC47基因的表达模式分析
-
NAC转录因子家族是一类植物特异性调控蛋白,广泛存在于各种植物物种中,从模式植物拟南芥到重要作物如水稻和小麦(Triticum aestivum L.),通常超过100个成员,在植物生长发育、胁迫应答、代谢途径等多种生物学过程中发挥不可或缺的作用[23, 26-27]。本研究基于课题组前期的蛋白互做筛选,成功从蒺藜苜蓿MtUFO上游转录因子中鉴定出MtNAC47基因。前期研究已证明MtUFO在蒺藜苜蓿叶片和花器官发育中发挥重要作用,因此为探究MtNAC47基因在蒺藜苜蓿基因表达调控中的具体功能,克隆了蒺藜苜蓿中的MtNAC47基因,对其表达模式、转录自激活活性及亚细胞定位等进行分析,旨在为揭示该转录因子的功能和作用机制提供理论基础。
进化树分析发现,MtNAC47与红三叶TpNAC47亲缘关系最近,它们可能由共同的祖先进化而来,并具有相似的生物学功能;多序列比对显示MtNAC47蛋白与毒羊豆、蓖麻、红三叶等同源蛋白相似度较高,且具有相同的NAM保守结构域,NAM结构域是NAC转录因子家族的一个标志性特征,它与转录激活、植物生长调节以及逆境响应等生物学过程密切相关[28];基因结构的差异分析及蛋白保守基序分析表明,不同物种的NAC基因在转录调控、基因表达和功能上存在多样性,MtNAC47在进化过程中保持了较高的保守性,且在功能上具有相似性和较强的稳定性。亚细胞定位结果显示MtNAC47定位于细胞核和细胞质,与前人研究结果一致,说明MtNAC47主要在细胞核和细胞质中行使功能[29-30]。蛋白亲疏水性分析表明,MtNAC47含有多个亲水区和疏水区,这可能与其在植物细胞中的功能和定位密切相关。磷酸化位点预测结果显示,MtNAC47蛋白共含有43个磷酸化位点,其中丝氨酸磷酸化位点最多,发生磷酸化修饰的可能性最高,推测该蛋白可能通过调控这些特定的磷酸化位点在翻译后修饰其转录因子活性,影响下游基因的表达。转录因子通过与其他蛋白质的相互作用,直接或间接调控基因转录,在植物的应激反应、发育过程及其他重要生理过程中发挥着关键作用。本研究中的酵母自激活检测结果表明,MtNAC47具有自激活活性,这意味着MtNAC47能够在不依赖外部激活信号的情况下自行触发报告基因的表达。因此,MtNAC47不可直接应用于后续的酵母双杂交(Y2H)系统。
启动子预测有助于揭示基因表达的调控元件,为后续基因表达模式分析提供依据。通过对MtNAC47基因进行顺式作用元件分析,发现其具有低温胁迫响应元件以及GA、ABA等激素响应元件,因此对蒺藜苜蓿进行了相应的胁迫及激素处理。在SA、GA处理后的MtNAC47基因表达量为先上升后下降,这与水曲柳(Fraxinus mandshurica Rupr.) FmNAC1的表达模式相类似[31]。MeJA处理后的MtNAC47表达量呈显著下降趋势,与辣椒(Capsicum annuum L.)中的CaNAC064表达模式一致,推测MtNAC47在蒺藜苜蓿中可能通过相同或相似的方式来发挥作用[32]。
组织表达分析结果显示,MtNAC47基因在花中表达量最高,而在果中最低,表明该基因在蒺藜苜蓿中具有组织特异性。已有研究表明,NAC转录因子在水稻[20]、拟南芥[21]和玫瑰(Rosa rugosa Thunb.)[33]等植物中参与根系生长、花瓣细胞扩增以及开花时间的调控,证明了NAC转录因子在植物生长发育过程中通过冗余或非冗余的方式发挥重要作用。因此,MtNAC47在花中的高表达说明该基因可能与蒺藜苜蓿的花发育、花器官形成或花期调控密切相关。此外,有研究报道指出,NAC转录因子在冷胁迫下会被诱导,参与植物对冷胁迫的反应;并在低温胁迫下可能通过ABA信号途径与NAC转录因子的调控网络相互作用,共同调控植物的应激反应[12, 34]。本试验发现,在ABA和低温胁迫处理下,MtNAC47的表达量均表现为先下降后上升的趋势,变化趋势一致,说明ABA和低温胁迫均能诱导MtNAC47的表达。然而,MtNAC47是否通过ABA信号通路响应低温胁迫仍需通过后续实验进一步验证。
-
本研究成功克隆了蒺藜苜蓿MtNAC47基因,编码340个氨基酸,为非跨膜性蛋白,不具有信号肽,并且是一个亲水不稳定的碱性蛋白;MtNAC47在进化过程中具有高度保守性,亚细胞定位于细胞核和细胞质;该蛋白具有转录自激活活性,不能直接用于后续酵母双杂交实验。MtNAC47基因的表达量在SA、GA、MeJA、ABA激素诱导及低温胁迫下均有响应,在花组织中表达量最高,说明MtNAC47不仅在花器官发育中具有重要功能,还可能在植物对这些激素和逆境胁迫的应答过程中发挥调控作用。以上结果为进一步研究该基因的功能和机制提供了理论参考。
 DownLoad:
DownLoad: