





-
开放科学(资源服务)标识码(OSID):

-
RNA修饰是基因表达调控中的重要转录后调节方式,其中N6-甲基腺苷(m6A)是真核生物mRNA中最常见的化学修饰,显著影响修饰转录本的稳定性和翻译效率[1]。m6A修饰过程由3类蛋白协同调控,Writer甲基转移酶负责催化修饰添加,Eraser去甲基酶负责去除修饰,而阅读蛋白Reader识别并结合m6A位点,进而调控RNA代谢过程[2]。这一动态可逆的修饰机制构成了一种关键的转录后调控轴[3]。在植物中,m6A修饰被证实参与多个生物学过程,包括胚胎发生[4]、生长发育[5]、形态建成[6]、开花时间调控[5]、胁迫响应[5]以及作物产量形成[7-8]。YTH结构域结合蛋白(ECT/YTP)作为一类含有YT521-B同源结构域的RNA结合蛋白,在真核生物中被广泛认定为m6A阅读蛋白[9],能够识别m6A修饰并参与调控RNA的代谢及功能多样性[10]。该蛋白家族广泛存在于真核生物中,最早于小鼠中发现,随后在人类、酵母等多种物种中被鉴定[11]。哺乳动物中的YTP可分为3类:YTHDF1-3(胞质m6A阅读蛋白)、YTHDC1(核内蛋白,参与剪接调控)和YTHDC2(具解旋酶活性)。它们均具有一个高度保守的YTH结构域,该结构域由4~5个α螺旋与6个β链构成α/β折叠,其中β链形成β桶状结构,α螺旋则起稳定疏水核心的作用[12-14]。此外,YTHDC2还包含解旋酶结构域、ANK结构域和DUF1065结构域[13]。动物YTP被证明通过影响RNA剪接、稳定性和免疫原性,从而在免疫调节和抗肿瘤免疫中发挥重要作用[15]。
与其他真核生物相比,植物ECT家族成员更为丰富。拟南芥和水稻中分别鉴定出13和12个ECT基因,标志着植物ECT系统鉴定研究的开端[16]。苹果MhYTP基因家族包含15个成员[17]。MhYTP2被报道通过调控靶标mRNA m6A修饰水平、稳定性和翻译效率影响苹果对低温[18]、干旱[18]、低氮[19]和病原菌(白粉和炭疽)[20-21]的抗性。小麦中鉴定出39个TaYTP基因,其表达与胁迫应答及发育进程密切相关[22]。草莓[23]、番茄[24]等作物研究也进一步证实YTP在植株生长和果实发育中具有关键功能。
芒果(Mangifera indica L.)作为漆树科芒果属的常绿乔木,是一种广泛栽培的热带果树[25],其生长过程中常面临低温[26]、干旱[27]和病虫害[28]等多种胁迫,这些因素严重影响芒果的产量与品质,制约芒果产业发展。为应对胁迫,植物通过调控基因表达以激活抗逆机制[29]。因此,解析植物逆境应答的分子网络并挖掘关键基因,已成为作物遗传改良的重要研究方向。目前,芒果ECT基因家族尚未得到系统鉴定,其在生物与非生物胁迫中的功能亦有待深入探讨。本研究旨在对芒果ECT基因进行全基因组鉴定,分析其基因结构、启动子顺式作用元件,以及其编码蛋白的系统发育关系和保守结构域,探究该家族基因在不同胁迫条件下的表达模式,以期为芒果抗逆分子育种提供理论依据。
HTML
-
基于YTH保守结构域(Pfam PF04146)的HMM模型,利用TBtools软件搜索“四季蜜”芒果基因组数据库(非公开数据),获取含YTH结构域的CDS序列(去除内含子后),翻译为蛋白序列。通过CD-Search验证YTH结构域的存在,并用BioXM软件计算蛋白的等电点和分子量。
-
用MEGA11计算最优进化模型,采用最大似然法构建系统发育树,并使用Evolview网站(
https://www.evolgenius.info/evolview/#/treeview )可视化系统发育树。 -
将MiECT序列提交到MEME网站(
http://meme-suite.org/tools/meme ),得到保守蛋白质基序。利用TBtools分析基因结构、保守结构域和基序。 -
从“四季蜜”芒果基因库下载基因上游2 000 bp DNA序列,上传到Plantcare网站(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )进行基因启动子区顺式作用元件预测。分析结果通过TBtools进行可视化分析。 -
试验材料为“四季蜜”芒果,将芒果种子去掉种皮取出种胚,用清水水培种胚,每天更换清水。2周后长出幼苗并将其进行移栽,基质与蛭石比例为9∶1。培养2个月后选择长势均一的幼苗进行对应胁迫处理。
低温:将2月苗龄幼苗放入4 ℃培养箱中模拟低温胁迫,分别于0、6、12和48 h采集成熟叶片,立即置于液氮中冷冻,-80 ℃保存。
干旱:在充分灌溉后进行,浇水后记为0,分别于0、6、12和48 h采集成熟叶片,立即置于液氮中冷冻,-80 ℃保存。
炭疽:在相对湿度为100%的条件下,对花后100 d的果实分别接种胶孢炭疽菌(Colletotrichum gloeosporioides)并进行黑暗处理,分别于0、6、12、48、72和96 h采集果实组织,立即置于液氮中冷冻,-80 ℃保存。
水泡病(Blister)和溃败病(Internal flesh breakdown)是由于果实中矿质元素不平衡所引起的生理性病害[30-31],样品采自广西大学标本园,液氮速冻,保存于-80 ℃冰箱。
采用FastPure Universal Plant Total RNA Isolation Kit试剂盒提取植物总RNA(诺唯赞生物),所有耗材均使用DEPC酶处理并高温灭菌。采用Hiscript Ⅳ All-in-one Ultra RT SuperMix for qPCR试剂进行逆转录(诺唯赞生物)。荧光实时定量PCR(RT-qPCR)染料为ChamQ SYBR qPCR master Mixture(诺唯赞生物),以芒果持家基因Actin为内参基因,用primer3 Plus网站设计荧光定量PCR引物,引物序列见表 1,试验重复3次。
-
采用Excel表格对数据进行初步汇总整理。利用IBM SPSS Statistics 22.0软件对各指标数据进行单因素方差分析,因素不同水平间差异显著性采用Tukey's检验(设置0.05显著性水平),用字母标记法进行显著性标注。利用Graphpad Prism 9.5制图。
1.1. 芒果ECT蛋白的筛选与特性研究
1.2. 系统进化分析
1.3. 蛋白保守基序、保守结构域和基因结构分析
1.4. 基因启动子区顺式作用元件分析
1.5. MiECT基因表达模式分析
1.6. 数据统计与分析
-
在“四季蜜”芒果基因库中共鉴定到12个ECT基因,并预测其编码蛋白的理化性质,如表 2所示。在这些ECT蛋白中,MiECT8的氨基酸序列最长,为1 083个氨基酸,分子量约为119.6 kDa;而MiECT12最短,仅含203个氨基酸,分子量约为23.1 kDa。这些蛋白的等电点分布范围为5.12(MiECT10)至8.42(MiECT12)。
-
为探究ECT家族成员间的进化关系,用芒果、拟南芥、苹果、番茄、人类和甜橙这6个物种共66条ECT蛋白序列构建了系统发育树,如图 1所示。进化分析表明,芒果ECT蛋白可划分为YTHDF和YTHDC两个亚家族。其中,仅MiECT12和MiCPSF30归属于YTHDC1亚类,这一分类特征与拟南芥中11个ECT蛋白属于YTHDF亚家族、2个属于YTHDC亚家族的情况类似。所有分析的植物ECT蛋白中均未发现与人类HsYTHDC2直系同源的蛋白。进一步亲缘关系分析显示,芒果ECT与其他物种存在若干直系同源关系,尤其与甜橙的ECT(YTH)基因亲缘较近,如CsYTH4与MiECT6、CsYTH2与MiECT8、CsYTH7与MiECT4等。此外,芒果也与拟南芥(如AtECT5与MiECT9)和苹果(如MhYTP16与MiECT10)等物种的ECT基因表现出较近的进化关系。
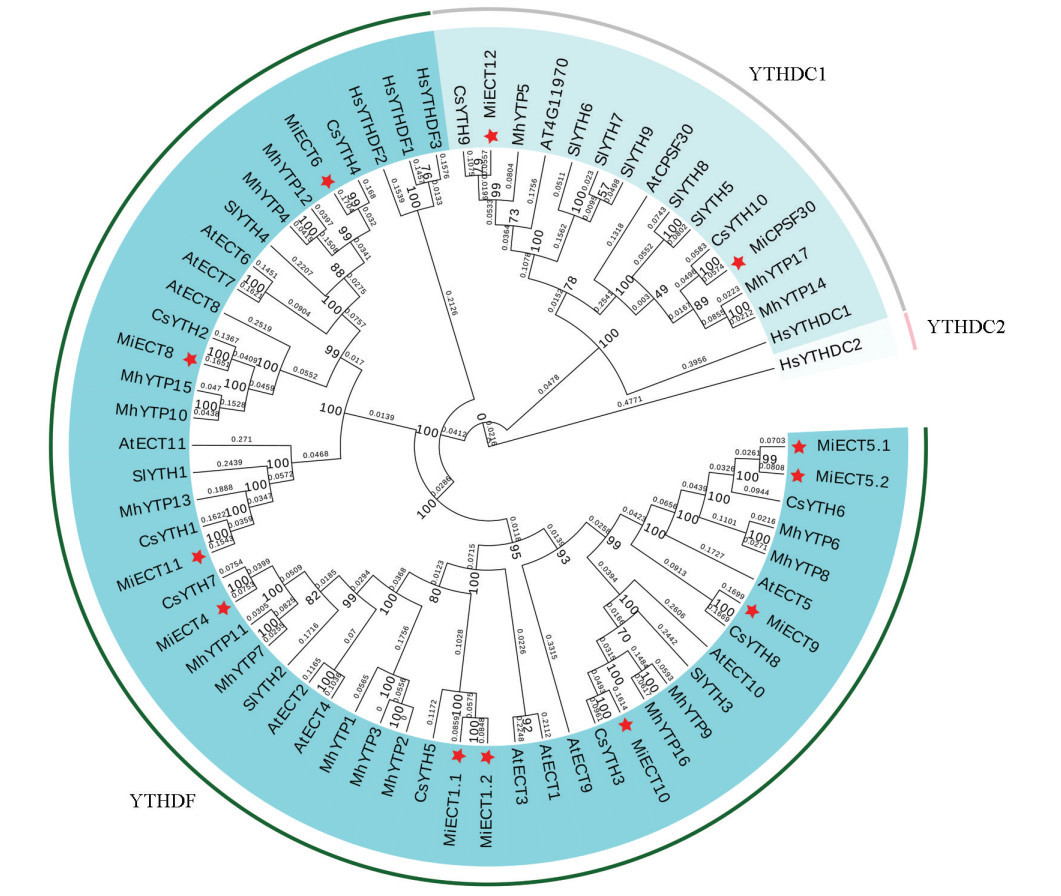
-
基序分析结果如图 2a所示,在MiECT家族中,除MiECT12和MiCPSF30,所有成员均包含6个按相同顺序排列的基序,即Motif5、Motif15、Motif2、Motif4、Motif1和Motif3。
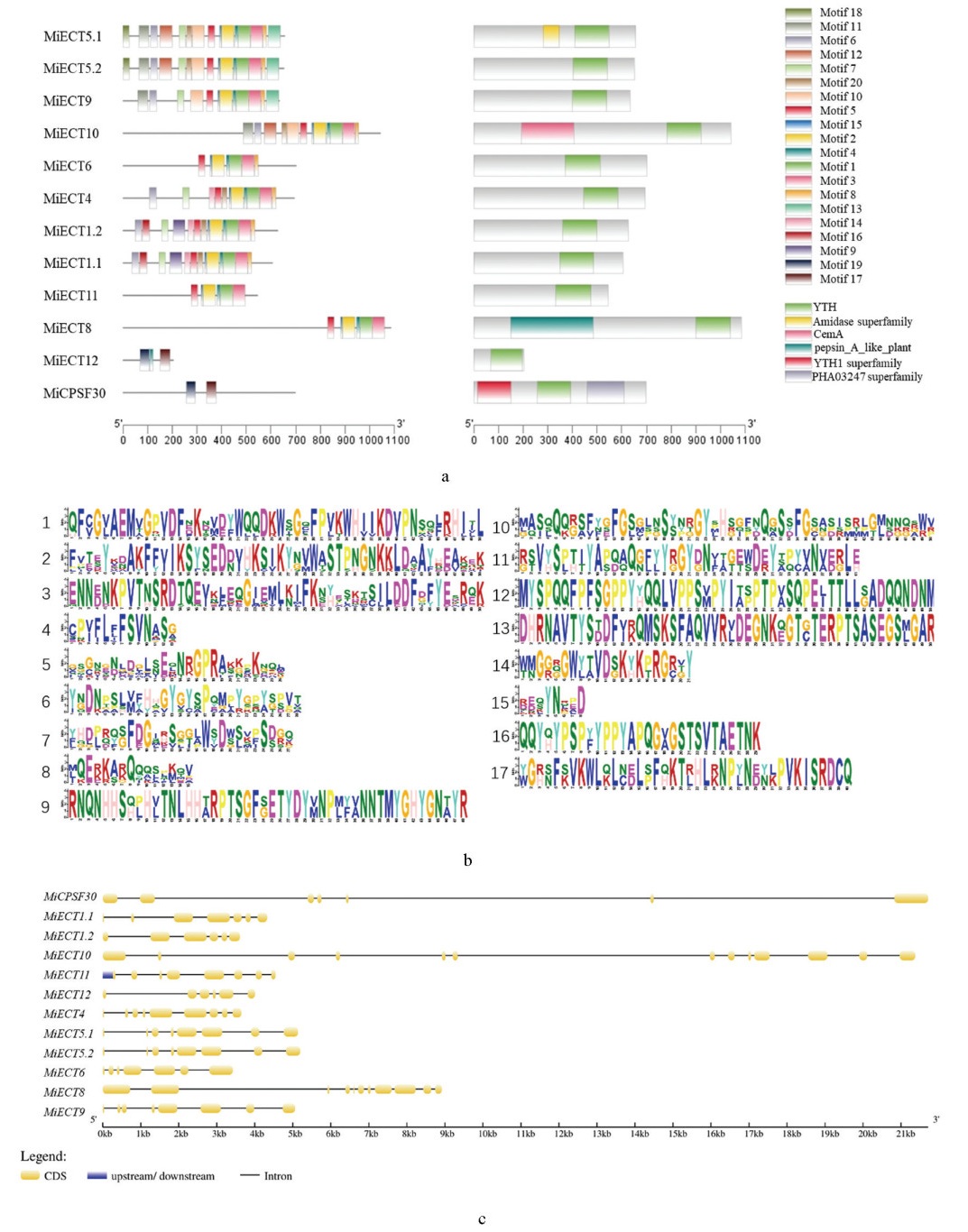
MiCPSF30所含基序数量最少,仅包括Motif19和Motif17,比MiECT12少一个基序。保守结构域分析表明所有MiECT蛋白均包含典型的YTH结构域,但其在蛋白质中的位置存在亚家族特异性,YTHDF亚家族的YTH结构域更靠近C端,而YTHDC1亚家族的MiECT12和MiCPSF30的YTH结构域则位于蛋白质中部。除YTH结构域外,未发现其他保守结构域(图 2b)。这些高度一致的基序排列和结构域分布模式表明,MiECT在进化上具有高度保守性。通过对比MiECT基因的mRNA序列与基因组序列,并利用TBtools软件绘制了12个MiECT基因的外显子-内含子分布图。分析显示,所有MiECT家族成员均含有不少于5个内含子,且大部分基因在外显子和内含子的大小及排列方式上存在显著差异,表明这些基因可能经历了独立的进化过程。值得注意的是,MiECT5.1与MiECT5.2,MiECT1.1与MiECT1.2表现出高度相似的基因结构(图 2c),提示它们可能来源于串联重复事件。
-
分析MiECT基因启动子顺式作用元件,结果如图 3所示,发现MiECT基因启动子区含有丰富的激素响应、胁迫响应及光响应等相关顺式作用元件,包括茉莉酸甲酯(MeJA)响应、脱落酸(ABA)响应、厌氧诱导响应、赤霉素响应、低温响应等元件。其中,参与厌氧诱导响应、茉莉酸甲酯(MeJA)响应和脱落酸响应的顺式作用元件几乎存在于所有MiECT基因中。
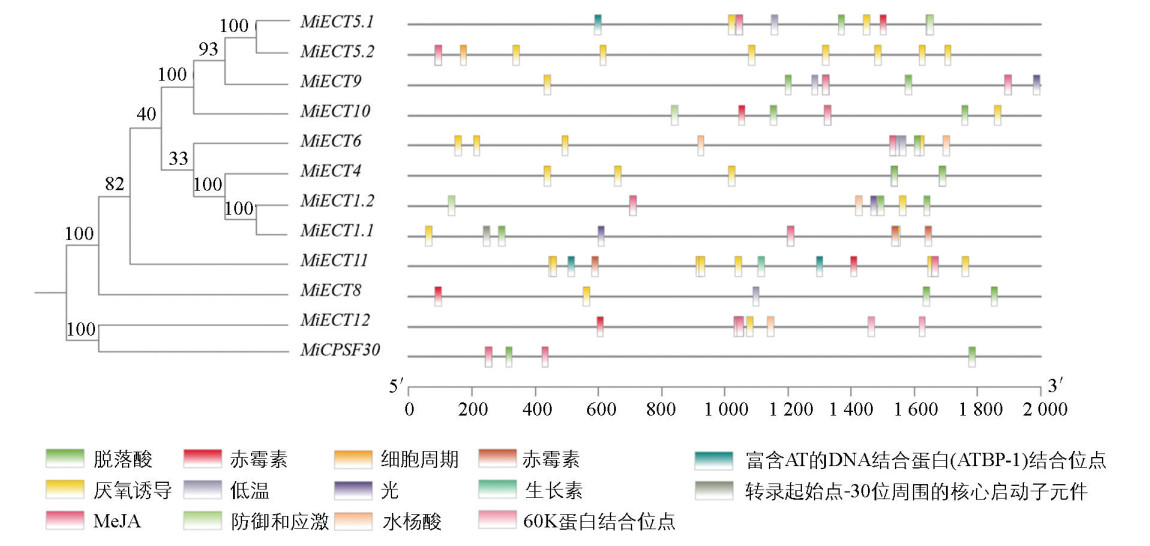
-
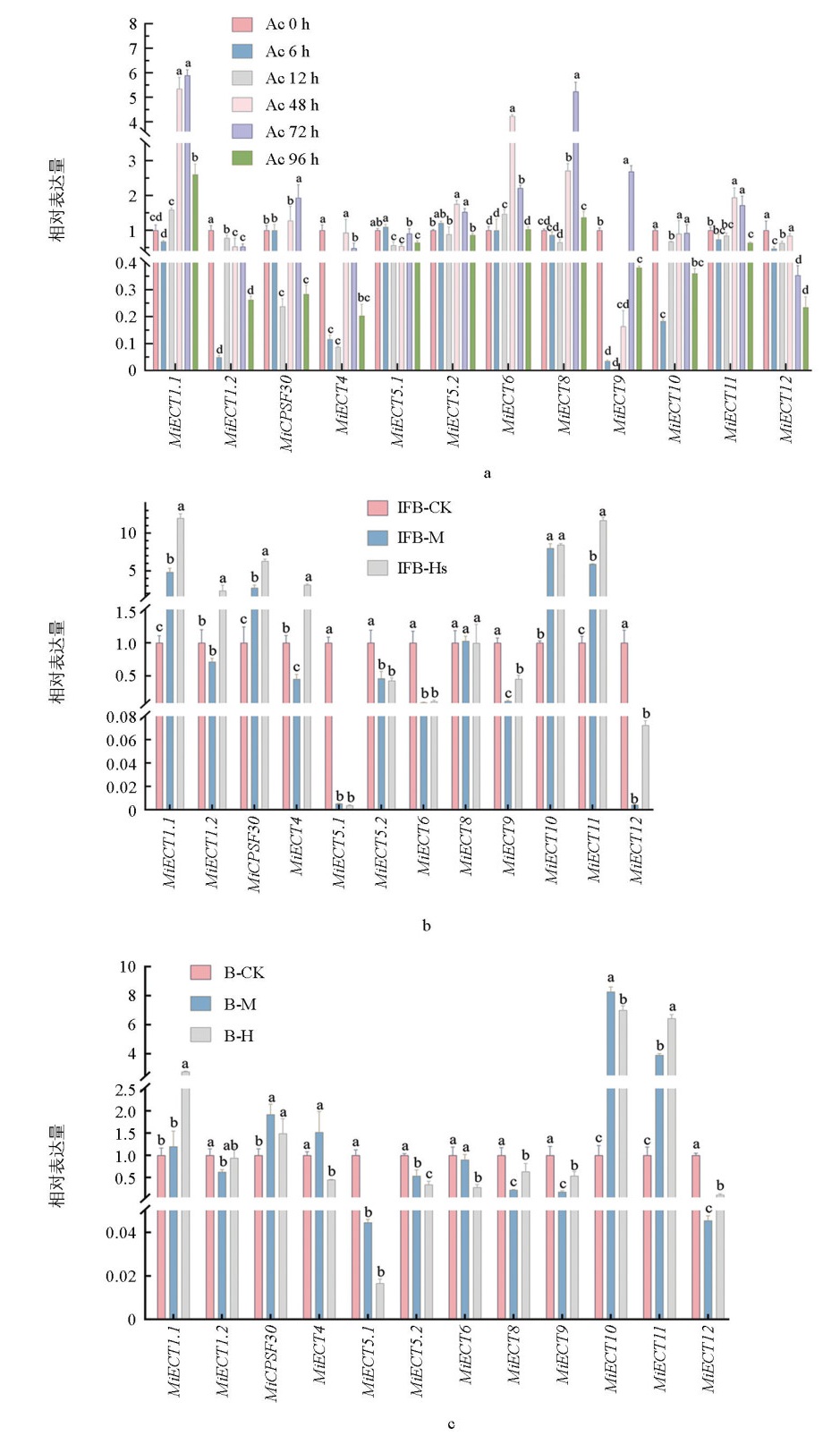
为探究MiECT基因在非生物中可能的生理功能,采用RT-qPCR技术首先分析了该家族基因在低温胁迫下的表达模式。从图 4可以看出,除MiCPSF30、MiECT9和MiECT10外,其他MiECT基因均受低温诱导显著表达。MiECT4和MiECT12表现出极为强烈的应答:MiECT4在胁迫6 h后的表达量急剧增至对照组的18倍;而MiECT12的反应稍晚,在12 h后表达量升至对照组的24倍(图 4a),表明它们在低温响应中可能扮演关键角色。
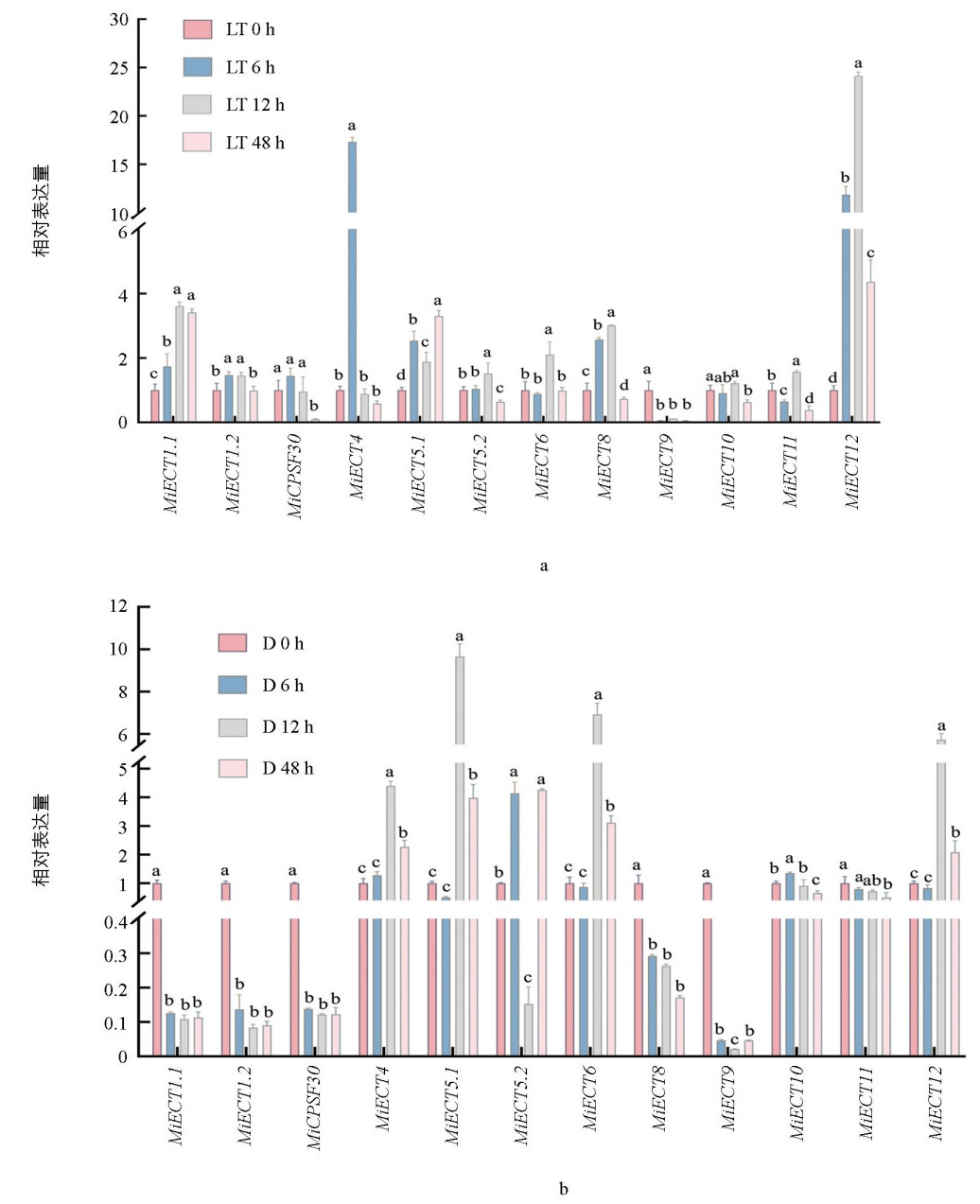
此外,分析了MiECT家族基因对干旱胁迫的响应。干旱处理显著抑制了MiECT1.1、MiECT1.2、MiCPSF30、MiECT8、MiECT9和MiECT11的表达;相反,MiECT4、MiECT5.1、MiECT5.2、MiECT6、MiECT10和MiECT12的表达则被显著诱导(图 4b)。
-
在炭疽病菌胁迫下,MiECT基因家族呈现出不同的表达模式:MiECT1.1、MiCPSF30、MiECT5.2、MiECT6、MiECT8、MiECT9和 MiECT11的表达显著上调;而 MiECT1.2、MiECT4、MiECT5.1、MiECT10和 MiECT12的表达则受到抑制(图 5a)。在溃败病发生后,MiECT基因的表达变化与病害严重程度呈现显著关联。其中,MiECT1.1、MiECT1.2、MiCPSF30、MiECT4、MiECT10和 MiECT11的表达水平与病情严重程度呈正相关;与之相反,MiECT5.1、MiECT5.2、MiECT6、MiECT9和 MiECT12的表达水平则与病情严重程度呈负相关。此外,MiECT8的表达未发生显著变化,表明其不响应此胁迫(图 5b)。在水泡病发生后,MiECT基因家族的表达出现分化:MiECT1.1、MiCPSF30、MiECT10和 MiECT11的表达被显著诱导;而 MiECT1.2、MiECT4、MiECT5.1、MiECT5.2、MiECT6、MiECT8、MiECT9和 MiECT12的表达则受到显著抑制(图 5c)。上述结果表明,MiECT基因家族成员在不同胁迫条件下具有特异性的表达调控模式,可能广泛参与芒果对逆境胁迫的响应过程,并与植物的抗逆机制密切相关。
2.1. 芒果MiECT基因家族的成员鉴定及其编码蛋白的理化性质分析
2.2. 芒果MiECT家族成员的系统进化分析
2.3. 芒果MiECT基因家族编码蛋白的保守基序、保守结构域及其基因结构分析
2.4. 芒果MiECT基因启动子区顺式作用元件分析
2.5. 芒果MiECT基因在非生物胁迫处理下的表达模式分析
2.6. 芒果MiECT基因在生物胁迫和生理性病害发生下的表达模式分析
-
MiECT蛋白作为m6A的主要阅读蛋白,在真核生物中具有高度保守性[32]。植物ECT家族成员数量普遍多于动物,例如拟南芥中有13个[33]、水稻12个[16]、苹果15个[18]、番茄9个[34]、甜橙10个[35],而普通小麦中更鉴定出39个ECT基因[22],均远多于人类的5个。本研究在芒果中鉴定到12个MiECT,其启动子含有多种激素与逆境响应元件,包括茉莉酸甲酯、赤霉素和低温响应元件等,表明该家族可能通过激素信号通路参与芒果的逆境应答机制。例如MiECT4启动子区域未发现典型的低温响应元件,但是存在脱落酸响应元件,其受低温处理诱导显著表达。已有研究表明,在水稻中,低温胁迫可通过ABRE顺式作用元件诱导ABA反应基因上调[36];而ABA作为一种关键的胁迫激素,在低温等渗透胁迫应答中起核心作用[37]。由此推测MiECT4在低温下的诱导表达可能与ABA信号通路有关。
多项研究表明,ECT基因在不同物种中广泛参与非生物胁迫响应,如低温、高温、干旱、盐和低氧等[38-39]。例如,AtECT12作为一个潜在m6A阅读蛋白,在盐和干旱胁迫中通过调节特定m6A修饰转录本的稳定性,增强植株抗逆性[40]。苹果MhYTP1和MhYTP2受ABA诱导,通过结合靶标RNA调控水分运输和气孔行为,增强抗旱性[18];MhYTP2还可通过结合并调控低温响应基因(MdRH20和MdGRP2),增强植株耐寒性[41]。番茄SlYTH8过量表达导致对冷胁迫更敏感,可能与表皮毛形态变化相关[42]。甜橙CsYTH2和CsYTH4受多种非生物胁迫及ABA诱导,表明其通过ABA信号通路在多重胁迫应答中发挥功能[35]。这些结果共同说明ECT基因在植物抗逆过程中具有重要作用。本研究系统分析了芒果MiECT基因在多种胁迫条件下的表达模式,结果显示该家族成员呈现多样化的表达响应。MiECT1.1、MiECT1.2、MiECT4、MiECT5.1、MiECT5.2、MiECT6、MiECT8和MiECT12响应低温胁迫显著上调表达;MiECT4、MiECT5.1、MiECT5.2、MiECT6、MiECT10和MiECT12受到干旱诱导显著上调表达;MiECT1.1、MiCPSF30、MiECT5.2、MiECT6、MiECT8、MiECT9和MiECT11受到炭疽病菌侵染显著上调表达;MiECT1.1、MiECT1.2、MiCPSF30、MiECT4、MiECT10和MiECT11的表达水平与溃败病发病程度呈正相关;MiECT1.1、MiCPSF30、MiECT10和MiECT11的表达被水泡病胁迫诱导显著上调表达。表明芒果MiECT家族可能通过类似的分子机制参与胁迫响应。
-
芒果中共鉴定出12个ECT家族成员,其中10个属于YTHDF亚家族,仅包含一个位于C末端的YTH结构域,未发现其他典型结构域;其余两个成员MiECT12和MiCPSF30属于YTHDC亚家族。MiECT家族在进化上具有高度保守性。表达谱分析表明,该家族成员广泛响应低温、干旱等非生物胁迫以及炭疽病、水泡病和溃败病等生物胁迫,但各成员在不同胁迫类型及处理时间点表现出高度差异化的表达模式。这些结果为进一步解析MiECT基因的功能机制奠定了重要基础。
 DownLoad:
DownLoad: