





-
开放科学(资源服务)标识码(OSID):

-
黄萎病是一种主要由大丽轮枝菌(Verticillium dahliae)侵染引起的土传维管束病害。黄萎病菌能够侵染马铃薯、茄子、番茄、向日葵和棉花等600多种植物,特别是双子叶植物几乎都能被侵染,给我国乃至世界农业生产造成严重的经济损失[1]。在我国,黄萎病对棉花生产的威胁尤为突出,据统计,棉花黄萎病年发病面积占植棉区50%以上,一般导致减产10%~30%,严重田块减产达80%甚至绝收,每年直接经济损失高达15亿~20亿元[2-4]。由于黄萎病病原菌可在土壤中存活数十年,且宿主范围广、传播途径多、变异频繁,因而防治难度大[5-7]。因棉花栽培中缺乏广谱持久抗性的抗原,加之难以防治,黄萎病被称为棉花的“癌症”[2]。因此,挖掘可利用的基因资源并阐明其抗病机制,可为棉花抗黄萎病分子育种提供理论支持。
HypSys(Hydroxyproline-rich Systemins,富含羟脯氨酸的系统素)是植物受机械损伤或病原菌侵染时激活免疫反应的信号分子之一,是植物自身合成的小分子肽,属于系统素(Systemin,Sys)亚家族,富含羟基脯氨酸(Hydroxyproline,Hyp),其成熟肽由18~20个氨基酸组成,来源于有N-末端分泌信号的前体蛋白肽[8]。HypSys的氨基酸在通过分泌系统加工后,多聚脯氨酸会经历羟基化,随后再糖基化[9-10]。这类系统肽通过结构修饰、信号转导及代谢调控在植物防御中发挥着重要作用[8-9, 11-13]。HypSys最初在研究烟草系统性创伤信号时被发现,之后陆续在番茄、马铃薯、甘薯等茄科和旋花科植物中发现了它的同源基因。HypSys通过介导细胞间或系统性的信号传递,协调植物的局部和整体性防御反应[14],它们之间没有明显的序列相似性,但都可以被创伤诱导,激活蛋白酶抑制剂、茉莉酸(JA)或乙烯(ET)信号通路以及昆虫防御的相关基因,进而影响昆虫的取食或消化功能,达到抗虫的效果[10, 12, 15]。烟草中组成型表达HypSys的前体基因TobpreproHypSys-A,可显著增强烟草对棉铃虫(Helicoverpa armigera)幼虫的抗性[16];过表达IbpreproHypSys的甘薯能显著抑制斜纹夜蛾(Spodoptera litura)幼虫的生长[17]。植物的HypSys也能激活植物防御素基因,或诱导病程相关蛋白基因的表达,或增强抗氧化酶活性,减轻病原菌侵染引起的氧化应激,进而在病原菌的防御中发挥作用[12, 15, 18-19]。烟草和拟南芥中组成型表达番茄系统素前体蛋白(Prosystemin,Pro-SYS)能提高烟草和拟南芥对死体营养型真菌灰霉菌(Botrytis cinerea)的抗性[20-21];外源施用来自番茄的HypSys可增强拟南芥对死体营养型真菌黄瓜棘孢菌(Plectosphaerella cucumerina)的抗性[22]。研究表明,HypSys在植物对死体营养型病原菌和病毒病的防御中具有重要作用[21-24]。
引起黄萎病的大丽轮枝菌为兼性营养型病原菌,植物能通过激活JA信号途径提高对黄萎病的防御能力[25]。HypSys也能激活JA信号通路,而HypSys是否影响植物对黄萎病菌的防御尚不清楚。为了阐明HypSys对黄萎病抗性的影响以及黄萎病菌侵染过程中HypSys的可能作用机制,本研究首先通过外施方法,阐明HypSys对黄萎病抗性的影响;其次利用组成型表达HypSys前体基因的转基因材料,研究HypSys能否稳定提高植物对黄萎病的抗性;再利用RT-qPCR(实时荧光定量PCR)方法检测防御信号途径标记基因表达水平的变化,分析HypSys影响植物黄萎病抗性的可能作用机制,以阐明其在植物抵抗黄萎病菌侵染中的作用及其作用机制。
HTML
-
植物材料:使用的植物材料分别为哥伦比亚生态型(Columbia ecotype)拟南芥、Sanmon烟草、本氏烟、陆地棉栽培种冀棉14号。黄萎病菌菌株:落叶型强致病力菌株V991。工程菌:农杆菌感受态细胞LBA4404和GV3101,大肠杆菌感受态细胞DH5α。
-
SlHypSys成熟肽(序列为GRHDSVLPPPSPKTDPII)[12],由南京强耀生物有限公司合成,用无菌双蒸水溶解,调整浓度至10 μmol/L。完全生长的拟南芥、本氏烟和棉花叶片用SlHypSys溶液浸润的吸水纸包裹离体叶片的叶柄,并于叶片表面均匀喷洒SlHypSys溶液进行处理,喷施水的处理为对照。外施处理试验重复进行3次,每次试验中,拟南芥、本氏烟草和棉花用SlHypSys和水处理叶片各30叶,所有处理的叶片均接种病原菌进行抗性分析。
-
取少许样品组织,利用Eastep® Super总RNA提取试剂盒(LS1040,Promoga),按说明书提供的操作程序提取RNA;然后利用TaKaRa的PrimeScriptTM RT reagent Kit with gDNA Eraser(RR047A)反转录试剂盒,按说明书提供的程序合成一链cDNA。
-
NCBI搜索NtHypSys和SlHypSys前体的编码序列(Gene ID:AY033148.1和AY292201),依据序列登录号NC_134094.1和NC_09080分别设计NtHypSys和SlHypSys前体基因PCR扩增引物(表 1),利用NtHypSys F和NtHypSys R引物对扩增NtHypSys前体基因的CDS(Coding sequence,编码序列)序列;SlHypSys F和SlHypSys R引物对扩增SlHypSys前体基因的CDS序列。
4周龄本氏烟和番茄叶片,于叶柄基部用镊子轻轻夹伤,损伤处理4~6 h,按1.3的方法提取RNA和合成cDNA,然后以cDNA作为模板,利用设计的引物(表 1)和Primes STAR MAX DNA Polymerase进行PCR扩增,扩增程序为98 ℃ 3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 10 s,35个循环;72 ℃ 5 min。扩增产物进行琼脂糖凝胶电泳,并利用胶回收试剂盒回收目的片段,然后按图 1的流程图构建NtHypSys和SlHypSys前体基因的过表达载体。
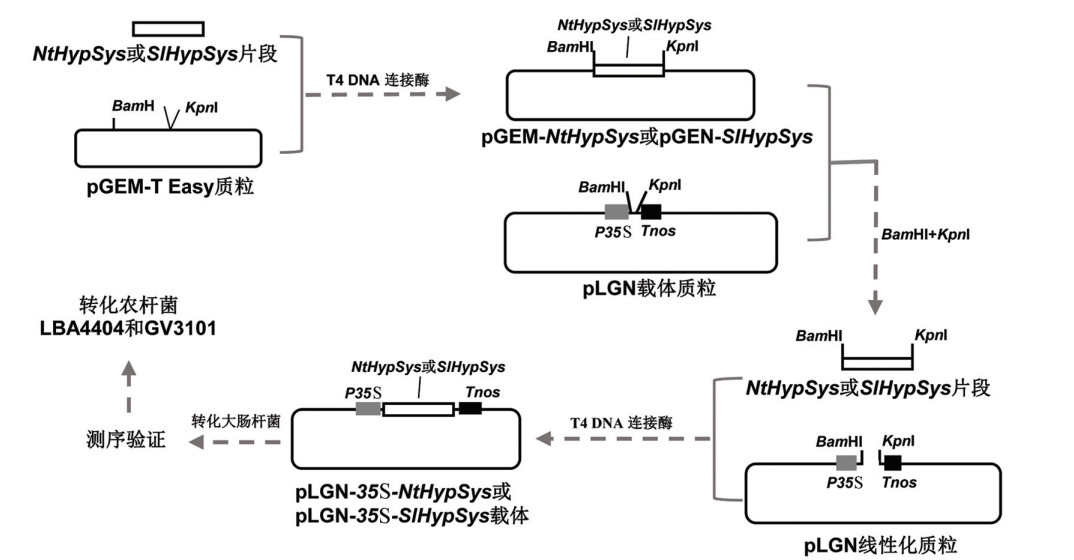
-
参照Clough等[26]农杆菌介导的浸花转化法转化拟南芥,并利用Harrison等[27]的方法筛选具有抗生素抗性的拟南芥幼苗。参照Zheng等[28]农杆菌介导的叶盘转化法,以Sanmon无菌烟草叶片为材料,转化烟草;参照Luo等[29]以冀棉14号无菌下胚轴为受体,农杆菌介导的棉花遗传转化法转化棉花。
-
参照Harrison等[27]的方法进行抗生素抗性筛选,获得具有卡那霉素抗性的植株。然后参照Jefferson[30]的方法,所有卡那霉素抗性植株均进行GUS(β-葡萄糖醛酸糖苷酶)组织化学染色检测,获得转基因植株。进一步对所有GUS染色阳性植株进行RT-qPCR检测,筛选表达水平显著提高的植株用于功能分析。RT-qPCR检测的内标基因为 AtACT3 (基因登录号AT3G53750),设置3个技术重复。RT-qPCR的20 μL反应体系包括:cDNA模板1 μL,基因上下游引物各1 μL,2×Taq SYBR Green qPCR Premix 10 μL,ddH2O 7 μL。扩增条件:95 ℃ 3 min;94 ℃ 10 s,57 ℃ 30 s,72 ℃ 30 s,共扩增40个循环。扩增完成后利用Gene study软件分析样本中目的基因的表达量,所有表达检测引物均用Primer Premier 5软件设计,序列见表 2。
转基因拟南芥纯合株系的筛选:每个转化子分单株收获种子,T2代开始,单株收获的种子取少许(100粒以上)水培至种子萌发产生胚根,然后进行GUS组织化学染色,若所有幼苗均能染成蓝色,则这个单株为纯合株系。如果T2代没有筛选到纯合的株系,T3代继续按此方法进行筛选。
-
转基因烟草再生过程中利用卡那霉素进行筛选,再生植株利用GUS组织化学染色进行鉴定,再利用RT-qPCR方法检测转基因的表达水平,筛选表达水平显著提高的转基因株系进行基因功能分析。
转基因烟草纯合转化子的筛选和表达水平检测方法同1.6.1。RT-qPCR检测的内标基因为NtGAPDH(LOC107817246),引物见表 2,设置3个技术重复。
-
转基因棉花再生过程中利用卡那霉素进行筛选,再生植株的GUS组织化学染色和RT-qPCR检测方法同1.6.1。RT-qPCR检测的内标基因为GhHIS3 (AF024716),引物序列见表 2,设置3个技术重复。
转基因棉花纯合株系的筛选:T2代转基因棉花开花前1 d,利用自交夹封闭花朵进行自交,挂牌标记开花日期,每株取开花后20 d的棉桃2个,剥取每粒种子的子叶,逐一进行GUS组织化学染色,若子叶能全部着蓝色,则这个单株及其后代为纯合株系。
-
外施SlHypSys拟南芥植株,接种黄萎病菌前(0 h)和接种后6、12、24、36、48、60、72 h的叶片;转基因和野生型(WT)拟南芥植株,接种黄萎病菌前(0 h)和接种后12、24、36、48 h的叶片,利用1.3的方法提取RNA和合成一链cDNA,然后以cDNA为模板,利用RT-qPCR方法检测SA信号途径的标记基因AtPR1 (AT2G14610)或AtNPR1 (AT1G64280)、JA信号途径标记基因AtPDF1.2 (AT5G44420)、NO信号途径标记基因AtNOS1 (AT3G47450)的表达水平。按照1.6方法进行RT-qPCR扩增,引物序列见表 3,设置3个技术重复。所有基因表达检测引物均参照TAIR数据库(
https://www.arabidopsis.org/ )中的基因序列,以AtACT3为内参基因,利用Primer Premier 5软件设计。 -
甘油保存的黄萎病菌V991菌株,挑取少许接种至PDA(马铃薯葡萄糖琼脂培养基)平板,26 ℃暗培养14 d,再刮取孢子接种至100 mL的PDB(马铃薯葡萄糖肉汤)培养基,26 ℃,200 r/min振荡培养7 d,4层纱布过滤去除菌丝,滤液5 000 r/min室温离心5 min,收集孢子,用去离子水调整孢子浓度为1.0×1010个/mL。
喷施SlHypSys的叶片表面晾干后,用吸头在棉花和本氏烟叶片主叶脉上轻轻挤压损伤2个点,拟南芥叶片在主叶脉中部损伤1个点,然后每个点接种5 μL孢子浓度为1.0×1010个/mL的黄萎病菌孢子悬浮液,保湿培养7 d后,按照5级统计叶片的病级和病情指数。SlHypSys处理离体叶片的抗病分析试验重复进行3次,每次处理和接种的拟南芥、棉花和烟草的叶片均为30叶。根据叶片的发病面积进行病级统计,分级标准为:0级:整个叶片均无病症;1级:叶片发病面积为(0~25%];2级:叶片发病面积为(25%~50%];3级:叶片发病面积为(50%~75%];4级:叶片发病面积超过75%[31]。
式中:I病情为叶片病情指数;N病叶为各级病叶数;G相应为相应级数;N总叶为调查总叶数;G最高为最高级别值。
-
1.8中培养的黄萎病菌液,用去离子水调整孢子浓度分别为1.0×107、1.0×108个/mL,用于转基因棉花、拟南芥和烟草植株的接种。
转基因拟南芥的抗病鉴定:按1.6.1的方法获得纯合的转基因拟南芥株系,T3代纯合的SlHypSys和NtHypSys转基因拟南芥,每转化子随机选取30株生长均匀、6~8叶的植株,利用灌根接种法,接种孢子浓度为1.0×108个/mL的黄萎病菌孢子液10 mL/株,然后12 h光照(26 ℃)、12 h黑暗(20 ℃)条件培养。接种14 d,按照5级统计法统计植株的病级,计算每个转化子的病情指数。T1代转基因拟南芥GUS染色分离的不着蓝色的植株为非转基因植株,这部分植株进行繁殖,用作抗病鉴定的对照,试验重复3次。
转基因烟草和棉花的抗病鉴定:按1.6.2和1.6.3的方法筛选转基因烟草和棉花的纯合株系,纯合T3代NtHypSys、SlHypSys转基因烟草或棉花幼苗,利用灌根接种法,棉花每株灌10 mL孢子浓度为1.0×107个/mL的黄萎病菌孢子悬浮液,烟草每株灌50 mL孢子浓度为1.0×108个/mL的黄萎病菌孢子悬浮液,接种后幼苗的培养条件为16 h光照(28 ℃),8 h黑暗(20 ℃)。接种14 d和20 d,分别统计棉花和烟草植株的病级和转化子的病情指数,试验重复3次。
植株病级的分级方法:根据叶片发病比率进行病级统计,分级标准[31-32]:0级:整个植株均无病症;1级:发病叶片占(0~25%];2级:发病叶片占(25%~50%];3级:发病叶片占(50%~75%];4级:发病叶片超过75%。植物对黄萎病的抗性用植株病情指数进行评价。
式中:I′病情为植株病情指数;N病株为各级病株数;N总株为调查总株数。
-
所有数据都以 x±s表示,用t检验分析对照和处理、转基因和非转基因植株间的差异显著性。
1.1. 植物材料与菌株
1.2. SlHypSys的合成和外施处理
1.3. RNA提取和cDNA的合成
1.4. NtHypSys和SlHypSys前体基因过表达载体构建
1.5. 拟南芥、烟草和棉花的遗传转化
1.6. 转基因植株的分子鉴定及纯合株系的筛选
1.6.1. 转基因拟南芥的分子鉴定及纯合株系的筛选
1.6.2. 转基因烟草的分子鉴定及纯合株系的筛选
1.6.3. 转基因棉花的分子鉴定及纯合株系的筛选
1.7. 黄萎病菌侵染后,防御信号途径标记基因转录表达水平检测
1.8. SlHypSys处理的拟南芥、烟草和棉花叶片对黄萎病的抗性鉴定
1.9. 转基因拟南芥、烟草和棉花对黄萎病的抗性鉴定
1.10. 数据分析
-
为了分析HypSys在植物中防御黄萎病的作用,人工合成了番茄SlHypSys成熟肽,并对植物叶片进行喷施处理。植物叶片喷施SlHypSys后,定点接种黄萎病菌V991菌株。结果显示:接种菌株5 d,SlHypSys处理的拟南芥、烟草和棉花叶片失绿黄化或萎蔫的病症面积明显较水处理对照更小(图 2a)。病情指数统计结果显示:10 μmol/L SlHypSys处理的拟南芥、烟草和棉花的病情指数分别为36.78、27.22和18.75,相比水处理对照的51.97、54.25和44.64,分别下降29.23%、49.82%和58.00%,达到极显著水平(图 2b)。这些结果表明,外施SlHypSys能够提高拟南芥、烟草和棉花对黄萎病的抗性。
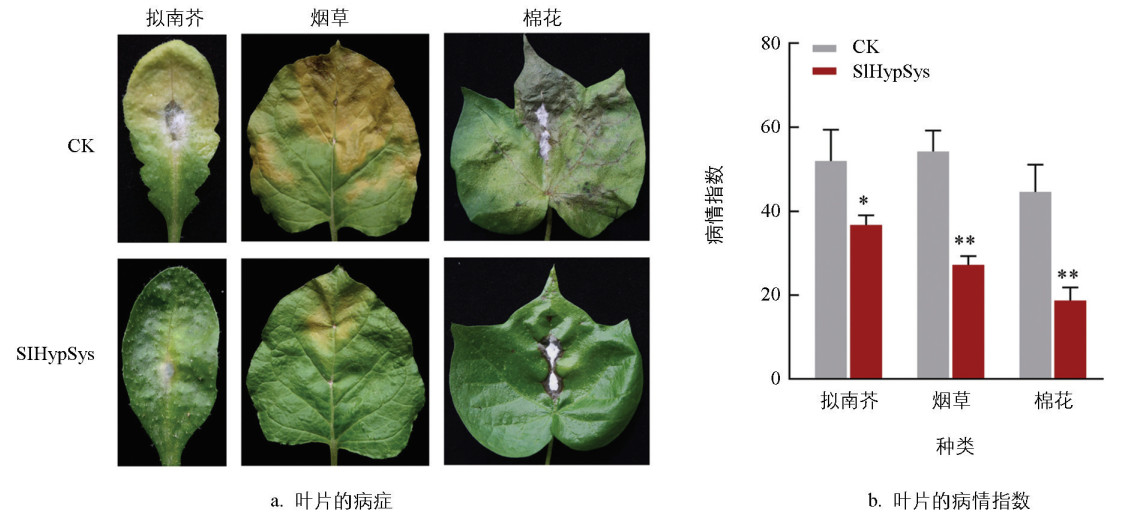
-
为了明确黄萎病菌侵染过程中SlHypSys影响植物黄萎病抗性的机制,利用RT-qPCR方法检测SlHypSys诱导防御信号途径标记基因的表达模式。结果显示:与喷施水的对照相比,黄萎病菌侵染过程中,喷施SlHypSys植株PR1的表达水平在接种后60 h大幅提高,接种12 h时提高了6.5倍(图 3a)。PDF1.2的表达水平从12 h开始提高,与喷施水的对照植株相比,喷施SlHypSys植株在接种72 h时PDF1.2的表达水平提高了38.3倍(图 3b)。NOS1的表达水平也有一定程度的提高,最高在接种后60 h达到3.5倍(图 3c)。这些结果表明,黄萎病菌侵染期间,外施SlHypSys可以提高SA、JA和NO信号途径标记基因PR1、PDF1.2和NOS1的表达水平。
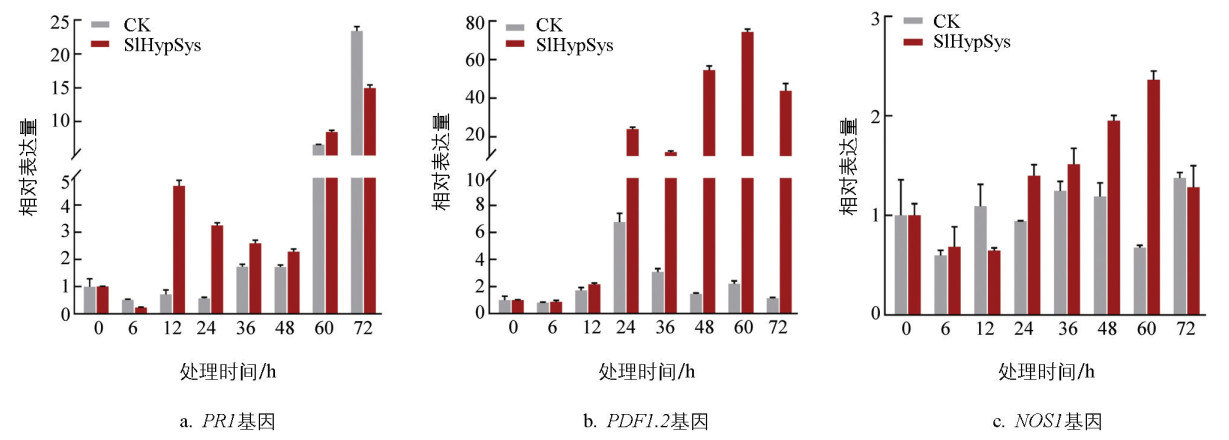
-
为进一步阐明HypSys在黄萎病菌侵染过程中的作用,利用PCR扩增法分别获得SlHypSys和NtHypSys前体的编码序列,构建其过表达的植物表达载体pLGN-35S-SlHypSys和pLGN-35S-NtHypSys(图 4a)。利用根癌农杆菌介导的遗传转化法将这两个载体分别转入拟南芥和烟草,经卡那霉素抗性筛选、GUS组织化学染色和RT-qPCR分析,最终获得5个烟草和11个拟南芥的NtHypSys转化子、6个烟草和12个拟南芥的SlHypSys转化子。
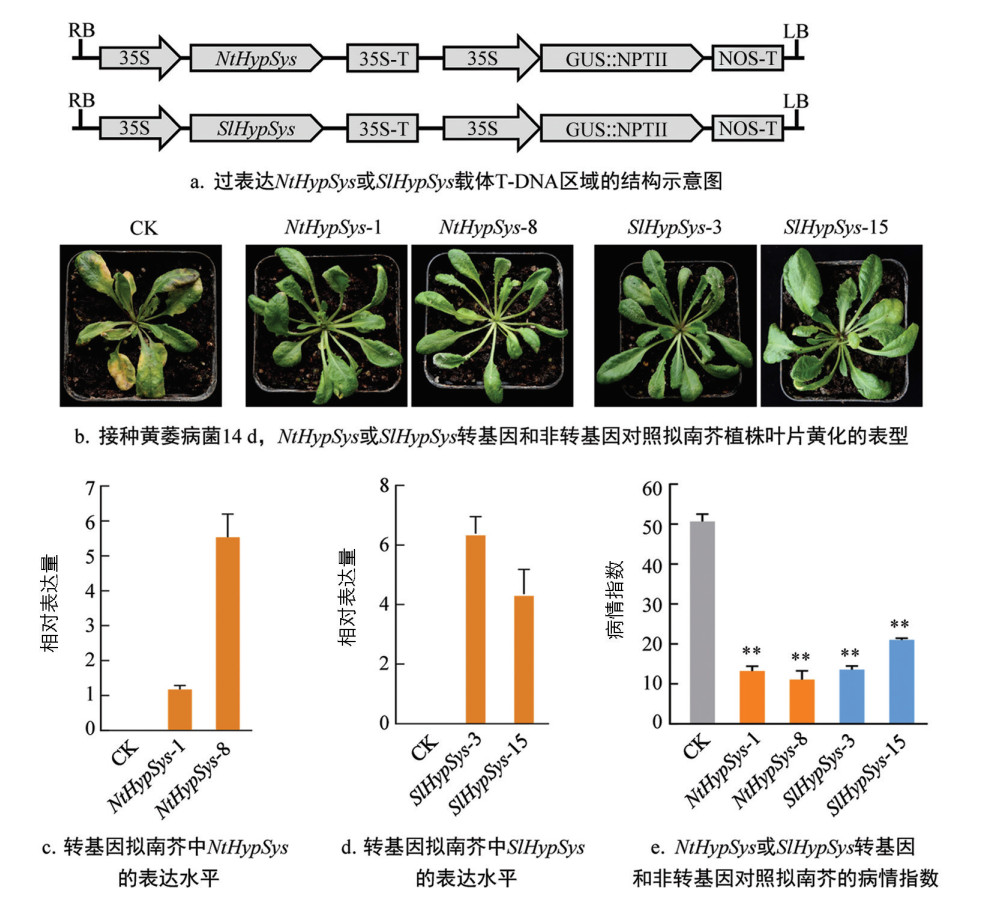
RT-qPCR结果显示,转基因拟南芥和烟草纯合株系中NtHypSys和SlHypSys都能稳定表达(图 4c-4d和图 5b-5c)。拟南芥浸根接种的抗病鉴定结果显示,接种黄萎病菌14 d,转基因株系中分离的非转基因对照植株叶片(CK)出现大面积失绿变黄等典型的黄萎病病症,但转基因植株仅底部少数叶片出现了病症(图 4b)。NtHypSys-1、NtHypSys-8、SlHypSys-3和SlHypSys-15转化子的病情指数分别为13.19、11.04、13.52和20.99,与非转基因对照(50.69)相比,转基因株系的病情指数极显著降低,降低比率分别为73.98%、78.22%、73.33%和58.59%(图 4e)。转基因烟草抗病鉴定结果显示,转基因株系中分离的非转基因对照植株(CK)多数叶片出现失绿变黄的黄萎病病症,而转基因植株仅底部叶片变黄(图 5a)。NtHypSys-2、NtHypSys-14、SlHypSys-1和SlHypSys-4转化子的病情指数分别为9.87、13.07、9.11和15.43,极显著低于非转基因对照的50.74,病情指数下降比率分别为80.55%、74.24%、82.05%和69.60%(图 5d)。这些结果表明,过表达来自烟草和番茄的类系统素前体基因SlHypSys和NtHypSys能显著提高拟南芥和烟草对黄萎病的抗性。
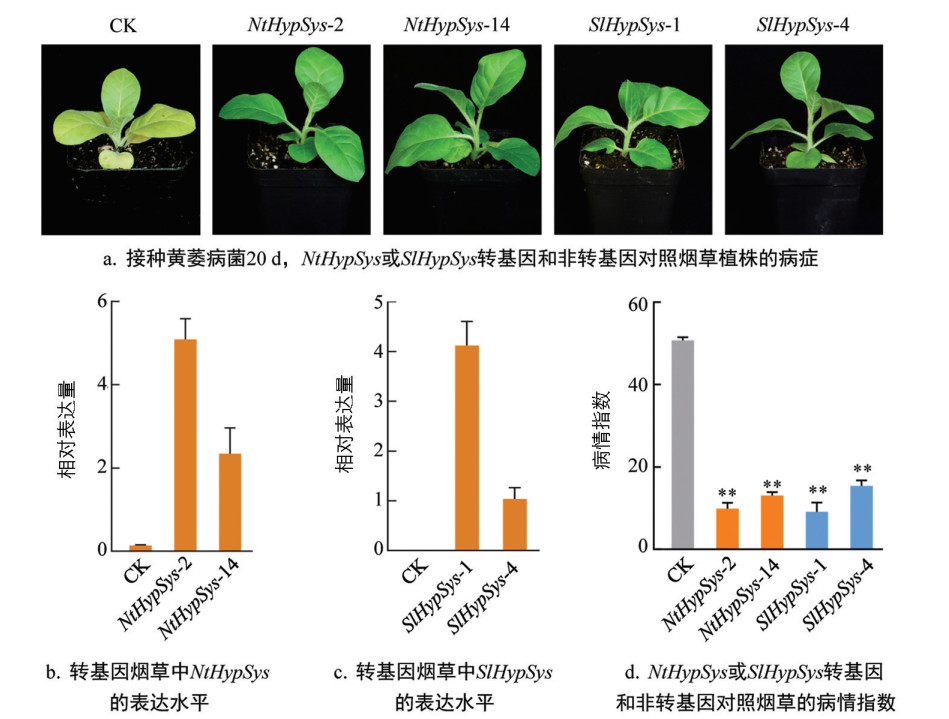
-
利用根癌农杆菌介导法将SlHypSys转入棉花,经卡那霉素抗性筛选和严格的分子鉴定后,获得独立转化子5个。灌根接种法接种黄萎病菌V991菌株14 d,转基因株系分离的非转基因对照植株叶片出现严重失绿的黄萎病病症,而转基因株系仅下部叶片出现了病症(图 6a)。RT-qPCR结果表明,SlHypSys在转基因棉花中得到有效表达(图 6b)。SlHypSys-1和SlHypSys-5转化子的病情指数分别为23.61和16.56,与非转基因对照(65.83)相比,转基因株系的病情指数分别下降64.13%和74.84%,达到极显著水平(图 6c)。综合表明,棉花中过表达SlHypSys前体基因能显著提高棉花对黄萎病的抗性。
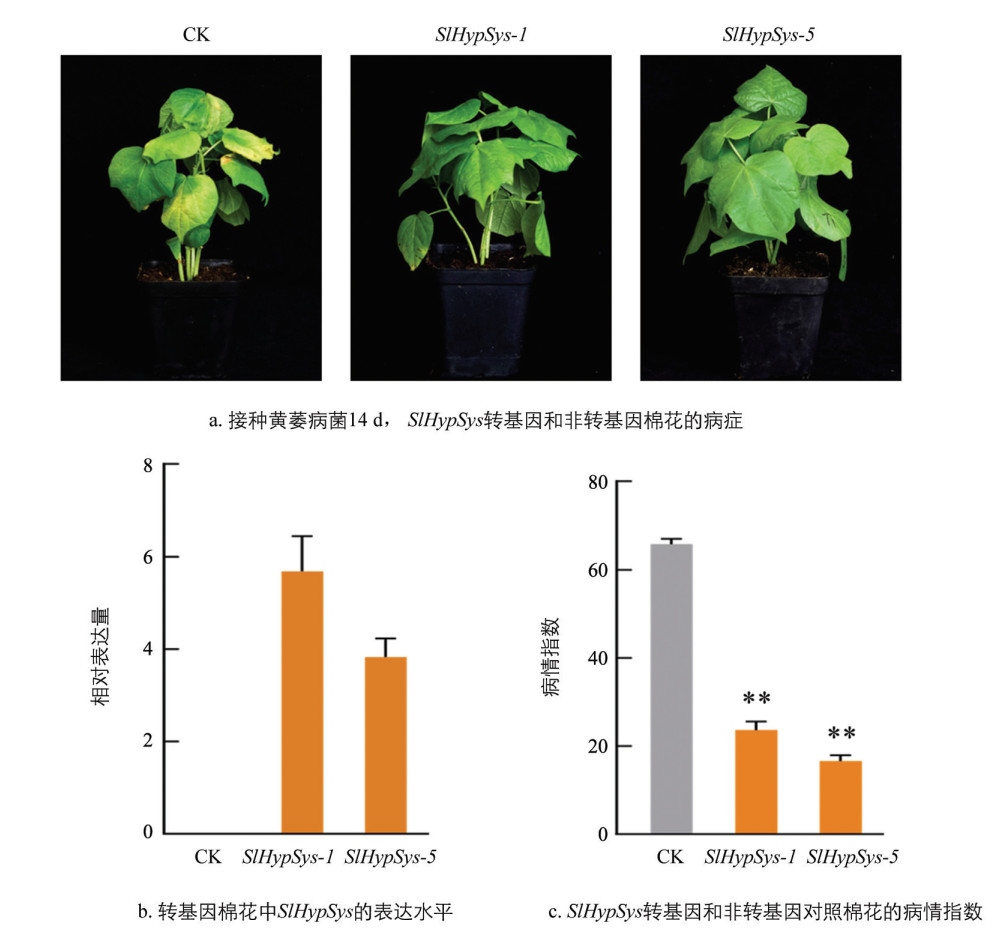
-
为进一步阐明HypSys能提高植物对黄萎病抗性的机制,利用RT-qPCR方法检测接种黄萎病菌后转基因拟南芥中SA、JA和NO信号途径标记基因的表达模式。结果显示,接种黄萎病菌后12 h和24 h,NtHypSys转基因拟南芥中RP1、NPR1、PDF1.2和NOS1的转录表达水平均明显高于对照;SlHypSys转基因拟南芥接种黄萎病菌后的12 h和48 h,RP1、NPR1、PDF1.2和NOS1的转录表达水平也高于对照(图 7)。PR1表达水平提高主要集中在接种黄萎病菌后36 h以内,而PDF1.2和NOS1表达水平提高持续时间更久。结果显示,拟南芥中异源表达SlHypSys和NtHypSys前体基因能有效提高防御反应信号途径标记基因PR1、NPR1、PDF1.2和NOS1的转录表达水平,表明NtHypSys和SlHypSys能通过激活植物的SA、JA和NO防御信号途径提高植物对黄萎病的抗性。
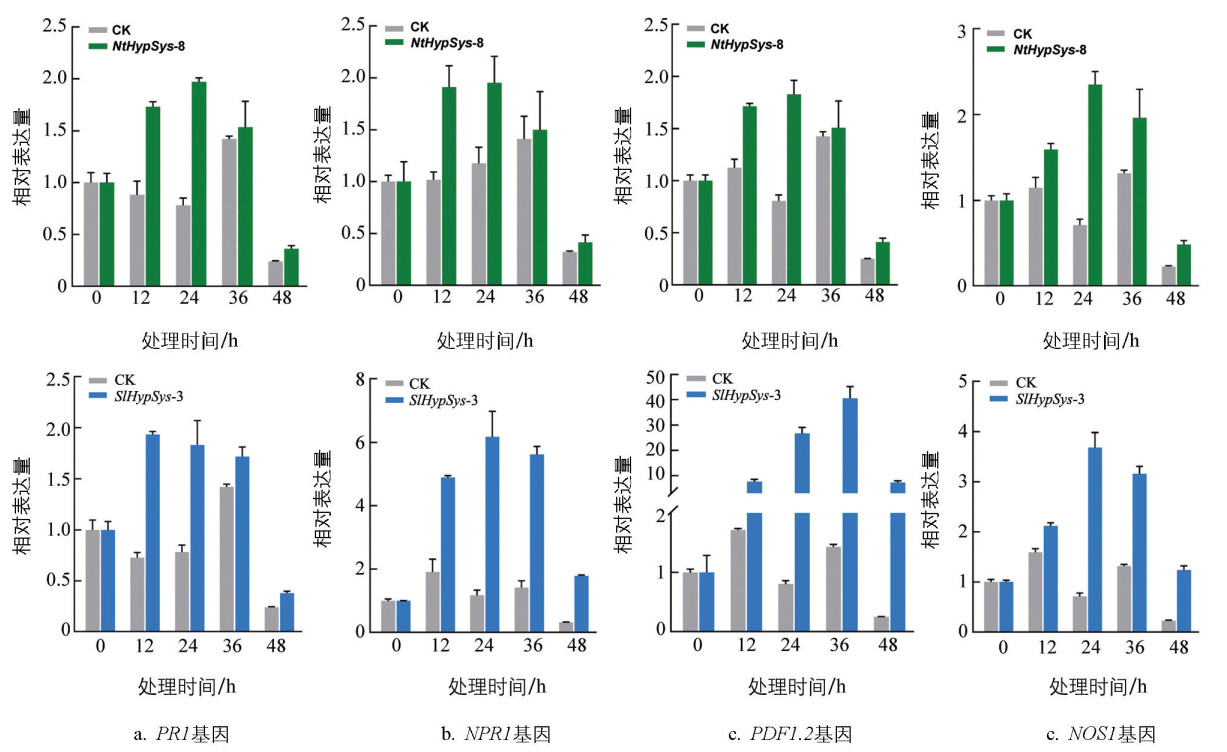
2.1. 外施番茄类系统肽SlHypSys能提高拟南芥、烟草和棉花对黄萎病的抗性
2.2. SlHypSys能提高防御信号途径标记基因的表达
2.3. 过表达SlHypSys和NtHypSys前体基因能提高拟南芥和烟草对黄萎病的抗性
2.4. 过表达SlHypSys前体基因能提高棉花对黄萎病的抗性
2.5. 黄萎病菌侵染过程中,SlHypSys和NtHypSys能促进防御信号途径标记基因的表达
-
Molisso等[33]研究表明,外施Systemin可以提高茄子和葡萄对灰葡萄孢的抗性。多位学者的研究表明,在番茄和拟南芥中组成型表达来自番茄的系统素前体基因Prosystemin,不仅能提高番茄对灰葡萄孢(B. cinerea)和链格孢菌(A. alternata)的抗性,还能提高拟南芥对灰葡萄孢和病毒病的抗性[21, 23-24]。Pastor-Fernández等[22]研究发现外施来自番茄的HypSys可以提高拟南芥对褐斑病菌(Plectosphaerella cucumerina)的抗性。上述研究表明,外源应用和内源增加HypSys或Sys的水平能提高植物对死体营养型病原菌的抗性。本研究发现,不管是外施还是通过转基因手段提高拟南芥、烟草和棉花内HypSys的水平,都可以有效提高其对黄萎病的抗性。黄萎病的致病菌大丽轮枝菌是一种土传维管束病原菌,具兼性营养特性[5],能通过植物的根或损伤部位侵染植物,然后在植物的维管组织内繁殖并扩展,加剧对植物的危害。HypSys或Sys等植物防御信号肽的前体蛋白或成熟肽在维管组织中积累,并在维管束内放大防御信号[14],因此推测HypSys可能通过在维管束内放大防御信号与黄萎病菌的侵染在空间上发生重叠,从而发挥一定的防御作用。
-
Pearce等[19]研究表明,来自矮牵牛的HypSys前体蛋白能特异性激活病原菌防御相关基因defensin1的表达;Bhattacharya等[12]利用外源供给的方式研究了马铃薯的HypSys(StHypSys)对病原菌防御相关基因表达的影响,结果表明,StHypSys能激活谷胱甘肽-S-转移酶GST3、水杨酸信号通路基因PAD4和NPR1等相关基因的表达;Pastor-Fernández等[22]研究表明,Systemin和HypSys增强植物的抗病性依赖于茉莉酸(JA)信号途径,并能在病原体侵染时增强由病原相关分子模式触发的免疫反应(PTI)。Bhattacharya等[12]则发现,HypSys能诱导ROS的产生,进而提高植物对病虫的抗性。在褐斑病菌侵染拟南芥时,系统素将激活MAPK信号途径,引发MPK3和MPK6的磷酸化,进而提高拟南芥对褐斑病的抗性[34]。本研究以拟南芥为材料,通过外施来自番茄的HypSys,发现在黄萎病菌侵染过程中,SlHypSys能激活SA、JA和NO信号途径标记基因PR1、PDF1.2和NOS1的转录表达水平(图 3),进一步在稳定遗传的转基因拟南芥中进行了类似的检测,结果显示,接种黄萎病菌后,NtHypSys和SlHypSys转基因拟南芥中PR1、NPR1、PDF1.2和NOS1的转录表达水平也都明显高于非转基因植株对照(图 7),表明番茄和烟草的类系统肽HypSys通过激活SA、JA和NO防御信号途径,进而提高植物对黄萎病的抗性。比较不同的研究结果发现,针对不同的病原菌,HypSys激活的防御反应存在一定的差异,但都是通过激活植物免疫相关的反应提高植物的抗病性。Pastor-Fernández等[34]的研究结果表明系统素和HypSys本身不具有直接的抑菌活性,本研究也发现SlHypSys没有抑菌活性,但外施应用SlHypSys和过表达NtHypSys和SlHypSys前体基因的转基因植株都能提高植物对黄萎病的抗性,研究结果证实,HypSys是通过激活植物免疫系统发挥保护作用。
-
本研究发现,不仅外源施加番茄的SlHypSys可以显著提高拟南芥、烟草和棉花对黄萎病的抗性,而且过表达NtHypsys和SlHypSys前体基因的转基因拟南芥、烟草和棉花纯合株系对黄萎病的抗性也显著提高。研究还表明,HypSys不仅在兼性营养型病原菌的防御中具有重要作用,还能跨物种提高植物对黄萎病的抗性。综合表明,来自茄科植物的外源HypSys能够被分类学上远缘的植物(如棉花和拟南芥)感知,受病原菌诱导时激活其防御信号途径,进而对兼性营养型病原真菌黄萎病菌(V. dahliae)产生抗性。该研究结果为HypSys在植物抗病育种中的应用提供了重要依据。
 DownLoad:
DownLoad: