




-
开放科学(资源服务)标志码(OSID):

-
玉米蚜(Rhopalosiphum maidis)属半翅目(Hemiptera)、蚜科(Aphididae)、缢管蚜属(Rhopalosiphum)昆虫,是禾谷类作物的重要害虫,主要危害玉米、高粱、谷子和小麦等,寄主超过了182种[1-3]. 玉米蚜具有繁殖能力强、世代周期短以及能够快速适应环境等特点,很容易大面积暴发[4]. 此外,玉米蚜在为害过程中由于分泌蜜露会导致煤污病的产生,同时还可能传播病毒病,不利于作物的生长发育,从而造成巨大的损失[5-6]. 防治玉米蚜的传统方法是化学防治,但玉米蚜作为典型的刺吸式口器吸食植物汁液的害虫,喜藏匿于叶片背面,一般药剂难以渗透进去. 其次,长期喷洒农药使蚜虫产生了极高的抗药性,同时容易对捕食性天敌造成为害,引发人畜中毒等问题,影响农业的可持续发展. 研究玉米蚜的绿色防治方法有利于助推农业高质量发展,因此,通过生物防治手段控制玉米蚜迫在眉睫.
玉米蚜常见的捕食性昆虫有食蚜蝇、寄生蜂、草蛉和瓢虫等[7],其中捕食性瓢虫是重要的天敌昆虫,在我国分布广泛[8]. 其中,黄斑盘瓢虫(Lemnia saucia)和六斑月瓢虫(Cheilomenes sexmaculata)在华南地区分布普遍,是常见的优势天敌瓢虫. 黄斑盘瓢虫的消长情况基本同多种蚜虫的为害高峰期相吻合[9],但近年来关于该瓢虫的研究主要是关注生活史特征、交配行为和繁殖特性等方面[10-14],对于该瓢虫对害虫的捕食潜力研究尚未见报道,一定程度上限制了黄斑盘瓢虫在生物防治中的应用. 六斑月瓢虫是广食性的昆虫[15],对于烟粉虱、蚜虫、粉蚧等害虫具有较强的防控潜力[16-17],目前已知捕食对象多达17种,包括10种蚜虫、2种粉虱、2种蓟马、2种木虱和1种夜蛾[17-18]. 因此,开展黄斑盘瓢虫和六斑月瓢虫对玉米蚜的捕食作用研究,对玉米蚜的科学防治具有重要意义.
黄斑盘瓢虫和六斑月瓢虫在农田中常常混合发生并控制害虫,但关于2种瓢虫对于玉米蚜的控害研究较少,尤其是黄斑盘瓢虫对玉米蚜的捕食作用还未见报道. 因此,本研究拟探讨黄斑盘瓢虫和六斑月瓢虫捕食玉米蚜的功能反应、寻找效应、种内干扰效应和自身密度干扰效应,进而明确黄斑盘瓢虫和六斑月瓢虫对玉米蚜虫的生物防治潜力,从而科学利用天敌昆虫防治玉米蚜虫.
HTML
-
黄斑盘瓢虫、六斑月瓢虫和玉米蚜均采自南宁师范大学武鸣校区校外的玉米田(108°17′E,23°10′N),瓢虫种群采集数量大于100头,在实验室条件下以玉米蚜饲养稳定种群,玉米蚜则长期使用玉米苗进行饲养. 选择羽化后10 d左右的黄斑盘瓢虫和六斑月瓢虫雌成虫,对生长情况较为一致的玉米蚜无翅成虫进行室内捕食作用试验,试验条件为温度(25±1) ℃,湿度(70±5)%,光周期为14L∶10D.
-
LRH-800C-GSIE智能人工气候箱(广东泰宏君科学仪器股份有限公司),直径为3.5 cm和9 cm的塑料培养皿,规格为25 cm×25 cm×25 cm、40 cm×40 cm×54 cm的养虫笼,马克笔和扫虫笔等工具.
-
黄斑盘瓢虫和六斑月瓢虫雌成虫饥饿处理24 h后进行试验. 在直径为9 cm的培养皿中放入一片新鲜的玉米叶片,挑选生长情况基本一致的玉米蚜成虫. 设置30、60、90、120、150头/皿5个玉米蚜密度梯度,每个密度下接入瓢虫雌成虫1头,24 h后观察并记录培养皿内剩余的玉米蚜数量. 每处理重复7次.
-
在直径为9 cm的培养皿中放入一片新鲜的玉米叶片,按照瓢虫雌成虫∶玉米蚜成虫=1∶30的比例,即将黄斑盘瓢虫和六斑月瓢虫雌成虫的密度设置为1、2、3、4、5头/皿,分别与30、60、90、120、150头/皿的玉米蚜成虫组合,24 h后统计瓢虫的取食量,每个处理重复7次.
-
在9 cm的培养皿中放入一片新鲜的玉米叶片,设置玉米蚜成虫数量为150头,分别测定瓢虫数为1、2、3、4、5头/皿时的捕食量,24 h后统计玉米蚜的存活数量. 每个处理设置7个重复.
-
采用逻辑回归分析玉米蚜被捕食量和起始量之间的关系,从而确定黄斑盘瓢虫和六斑月瓢虫成虫的功能反应类型. 将符合Holling Ⅱ型的数据利用圆盘方程Na=aTN/(1 + aTh N)进行拟合,式中Na为玉米蚜被捕食量,N表示玉米蚜密度,a为捕食者对玉米蚜的瞬时攻击率,T表示玉米蚜暴露给捕食者的时间,本试验中T为1 d,Th为捕食者处理1头蚜虫所耗时间[19].
利用公式S= a/(1 + aTh N)来计算寻找效应. 公式中,S为寻找效应,a为瞬时攻击率,Th为取食时间,N表示猎物数量[20].
采用Hassell模型公式评估种内干扰系数,其公式为:E=QP-m,其中,E表示模拟中的平均捕食率,Q为搜索常数,P表示天敌密度,m表示种内干扰系数[21].
根据公式I=(E1-EP)/E1计算分摊竞争强度I,式中E1为1头捕食者的捕食率;EP为捕食者密度为P头时的捕食率[22].
采用Watt模型方程进行拟合计算得出自身密度干扰效应结果,其方程为:A=aP-b,其中,A表示竞争条件下的平均捕食量,a表示常数(即无竞争条件下每头瓢虫的捕食量估计),P表示瓢虫密度,b表示竞争参数[23].
本试验数据采用Excel 2016和SPSS 20.0进行记录和统计分析,所有参数均采用最小二乘法进行计算.
1.1. 供试材料
1.1.1. 供试虫源
1.1.2. 主要设备和耗材
1.2. 试验方法
1.2.1. 黄斑盘瓢虫和六斑月瓢虫对玉米蚜的捕食功能反应和寻找效应
1.2.2. 种内干扰对黄斑盘瓢虫和六斑月瓢虫捕食率的影响
1.2.3. 黄斑盘瓢虫和六斑月瓢虫自身密度对捕食玉米蚜的干扰反应
1.3. 数据统计分析
-
黄斑盘瓢虫和六斑月瓢虫雌成虫对玉米蚜成虫的捕食量和捕食率见图 1. 2种瓢虫对玉米蚜的捕食量随着玉米蚜密度的增大而逐渐增大,当蚜虫数量增加到一定程度时,捕食量的增加速度变缓慢,捕食率下降. 经卡方检验,卡方值均大于0.05(表 1),意味着瓢虫对蚜虫的实际捕食量和理论捕食量间无显著差异,说明2种瓢虫对玉米蚜的捕食功能反应符合Holling Ⅱ型. 2种瓢虫在玉米蚜相同的密度梯度下取食量对比无显著差异. 在猎物密度为30头/皿时,黄斑盘瓢虫和六斑月瓢虫对玉米蚜的捕食率最大,分别为89.0%和98.1%;其次为85.0%和85.6%,即黄斑盘瓢虫和六斑月瓢虫分别在猎物密度为60头/皿和90头/皿时的捕食率(图 1).
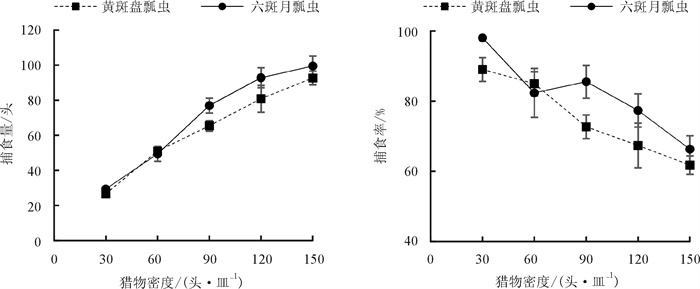
由表 1可知,黄斑盘瓢虫的瞬时攻击率(a)比六斑月瓢虫的低,二者分别为1.003 6和1.084 8;同时黄斑盘瓢虫对玉米蚜的处理时间(Th)也较长,为0.003 9 d,六斑月瓢虫的处理时间则为0.003 6 d;黄斑盘瓢虫和六斑月瓢虫对玉米蚜捕食作用的a/Th值分别为257.3和301.3,日最大捕食量分别为256.4头和277.8头.
-
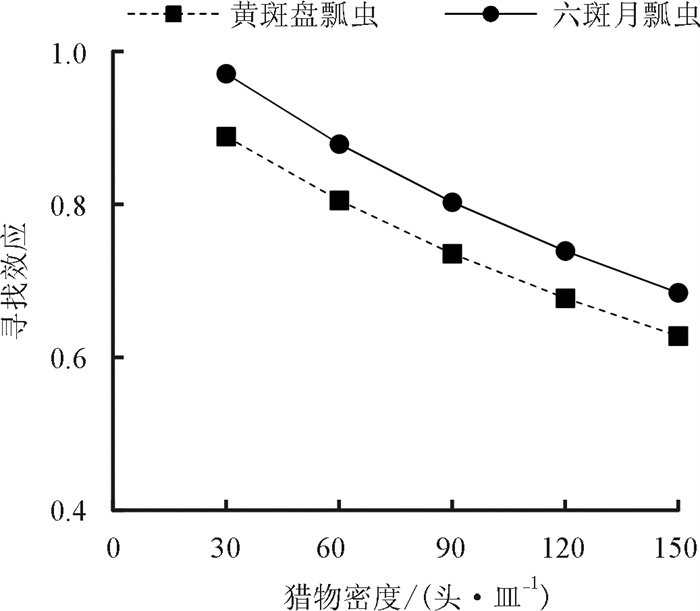
黄斑盘瓢虫成虫和六斑月瓢虫成虫对玉米蚜的寻找效应均随着玉米蚜密度的增加而逐渐下降,寻找效应方程分别为S=1.003 6/(1+0.003 9N)和S=1.084 8/(1+0.003 9N). 当猎物密度相同时,六斑月瓢虫对玉米蚜的寻找效应大于黄斑盘瓢虫的寻找效应(图 2).
-
黄斑盘瓢虫和六斑月瓢虫捕食玉米蚜的干扰系数(m)分别为0.963 9和0.999 9. 伴随着2种瓢虫密度的增加,一定空间内平均每头瓢虫的捕食量无显著性差异(黄斑盘瓢虫F=2.421,p=0.071;六斑月瓢虫F=0.856,p=0.502),但平均捕食率下降. 2种瓢虫的分摊竞争强度伴随着瓢虫密度的增加而增加,当瓢虫密度为5头/皿时,分摊竞争强度最大,表明在该密度条件下2种瓢虫的种内干扰作用最大(表 2).
-
2种瓢虫在一定空间和猎物密度不变的情况下,平均捕食量随自身密度的增加而下降(表 3),表明2种瓢虫捕食时存在种内干扰作用. 用Watt模型进行拟合,拟合方程、常数和相关系数等如表 3所示. 结果显示,黄斑盘瓢虫和六斑月瓢虫捕食玉米蚜的竞争参数分别为0.814 5和0.831 9. 根据Watt模型可知,随着2种瓢虫密度的增加,干扰作用增大. 将理论值和实际值经卡方检验,卡方值分别为0.912 3和0.979 8,均大于0.05,差异不显著,说明该模型能较好地反映黄斑盘瓢虫和六斑月瓢虫自身密度对捕食玉米蚜的影响情况.
2.1. 黄斑盘瓢虫和六斑月瓢虫对玉米蚜的捕食功能反应
2.2. 黄斑盘瓢虫和六斑月瓢虫对玉米蚜的寻找效应
2.3. 黄斑盘瓢虫和六斑月瓢虫的种内干扰效应
2.4. 黄斑盘瓢虫和六斑月瓢虫自身密度对捕食量的干扰反应
-
生物防治是实施绿色防控的有效途径,而利用天敌昆虫是生物防治的核心手段[24]. 在天敌昆虫中,捕食性瓢虫被认为是农业生态系统中最有前景的生物防治贡献者[25]. 黄斑盘瓢虫和六斑月瓢虫是我国华南地区常见的捕食性天敌,主要以蚜虫为食,在害虫生物防治的应用中具有巨大的开发潜能[26-29]. 本研究通过测定黄斑盘瓢虫和六斑月瓢虫对玉米蚜的捕食作用,评价了2种瓢虫的防治潜力. 研究结果表明黄斑盘瓢虫和六斑月瓢虫雌成虫对玉米蚜成虫的捕食量符合Holling Ⅱ圆盘方程,日最大捕食量超过250头,表现出较强的捕食能力. 本研究结果可为田间玉米蚜的绿色防治提供理论参考.
研究发现七星瓢虫(Coccinella septempunctata)取食豌豆修尾蚜(Megoura crassicauda)、草地贪夜蛾(Spodoptera frugiperda)[24, 30],异色瓢虫(Harmonia axyridis)捕食豌豆蚜(Acyrthosiphon pisum)、牛角花齿蓟马(Odontothrips loti)、柴胡明蚜(Hyadaphis bupleuri)[31-32]、六斑月瓢虫取食茶蚜(Toxoptera aurantii)、豆蚜(Aphis craccivora)、黄胸蓟马(Thrips hawaiiensis)、柑橘木虱(Diaphorina citri)和草地贪夜蛾(Spodoptera frugiperda)等害虫[17, 33-36]的捕食量可用Holling Ⅱ圆盘方程进行拟合,与本研究的功能反应模型一致. 模型中的a/Th值是衡量天敌捕食能力的重要参数之一,a/Th值越大,表明天敌控制能力越强[37]. 本研究结果中黄斑盘瓢虫对玉米蚜捕食作用的a/Th值小于六斑月瓢虫的值,表明六斑月瓢虫对玉米蚜的捕食能力要强于黄斑盘瓢虫. 唐良德等[37]的研究发现六斑月瓢虫雌成虫对玉米蚜成虫的最大捕食量为217.39头,本研究结果显示最大捕食量为277.8头,高于他们的结果,究其原因可能与试验的光照时间不同有关. 寻找效应是指天敌对害虫攻击的一种行为效应,天敌捕食作用的大小与其本身的寻找力度有关[30],黄斑盘瓢虫和六斑月瓢虫对猎物的寻找效应呈负相关,这与李秋荣等[38]、李盼等[39]和于静亚等[40]研究瓢虫捕食其他农林害虫的结果相似. 天敌在捕食时往往存在干扰效应,该效应大大地影响了天敌的捕食效率[41]. 本研究发现当天敌和猎物比例不变的条件下,黄斑盘瓢虫和六斑月瓢虫的捕食量随着猎物密度的增加而增大,但平均捕食率下降,分摊竞争强度增大,说明黄斑盘瓢虫和六斑月瓢虫的种内干扰作用明显. 除此之外,在玉米蚜密度保持不变的情况下,2种瓢虫的平均捕食量均随着自身密度的增加而下降,说明这2种瓢虫自身密度同样会对捕食产生影响.
在本研究的功能反应模型中,当猎物密度为30头时,瓢虫捕食率最高,但害虫密度太低,会失去吸引天敌的条件[40]. 当猎物密度为150头时,捕食量最大,寻找猎物所需时间最少,但捕食率最低,控害效果欠佳. 因此对于黄斑盘瓢虫和六斑月瓢虫分别推荐1∶60和1∶90的益害比进行释放,在此比例下2种瓢虫的捕食率较高,寻找猎物所需时间也较短. 但由于瓢虫种内干扰作用明显,因此在田间实际的生物防治应用中,应避免扎堆释放,分散多次释放以减少种内干扰和自身密度的影响,充分发挥天敌瓢虫的控害效果. 由于本研究是在室内的人工气候箱中进行的,而在自然条件下,田间的生态环境更为复杂,天敌的捕食能力不仅受到猎物密度、自身密度以及种内干扰的影响,同时还可能受到天气条件、猎物种类、天敌种间竞争等因素的影响[38, 41]. 因此,多环境因素对黄斑盘瓢虫和六斑月瓢虫捕食能力的影响需进一步探讨. 同时,应尽可能地创造类似于自然环境的试验条件,以实现更准确、更客观以及更合理地来评价天敌对害虫的实际控制作用. 本文探讨了黄斑盘瓢虫和六斑月瓢虫成虫对玉米蚜的捕食作用,但2种瓢虫共同存在时对玉米蚜的捕食作用以及它们幼虫对玉米蚜的控制能力还需进行深入研究.
 DownLoad:
DownLoad: